Key Points
Repertoire analysis by microarray technology constitutes a powerful tool to evaluate immune system failure in PADs.

Abstract
Immune system failure in primary antibody deficiencies (PADs) has been linked to recurrent infections, autoimmunity, and cancer, yet clinical judgment is often based on the reactivity to a restricted panel of antigens. Previously, we demonstrated that the human repertoire of carbohydrate-specific immunoglobulin G (IgG) exhibits modular organization related to glycan epitope structure. The current study compares the glycan-specific IgG repertoires between different PAD entities. Distinct repertoire profiles with extensive qualitative glycan-recognition defects were observed, which are characterized by the common loss of Galα and GalNAc reactivity and disease-specific recognition of microbial antigens, self-antigens, and tumor-associated carbohydrate antigens. Antibody repertoire analysis may provide a useful tool to elucidate the degree and the clinical implications of immune system failure in individual patients.
Introduction
Inadequate humoral responses to carbohydrate structures represent a common feature of primary antibody deficiencies (PADs), despite the pathogenetic heterogeneity of these disorders, involving monogenetic, polygenetic, and still-unexplained defects.1,2 These disorders are associated with a plethora of clinical sequelae, including severe and recurrent infections, microbial dysbiosis, autoimmunity, allergy, granulomatous disease, and malignancy.1,3,4
The surface of all living cells is glycosylated, and the composition varies between cell type,5 individuals, and species.6 The prominent exposure of carbohydrate structures (glycans) on the surface of cells or bacterial capsules renders them accessible to antibodies, which facilitates the detection and elimination of pathogens or aberrant cells, as well as leads to adverse reactions in transfusion/transplantation procedures (blood groups antigens) or to the acute rejection of xenografts (eg, Galα structures). Because of the altered expression of biosynthetic enzymes, such as glycosyltransferases or glycosidases, the glycome of cells is often changed under pathological conditions, including cancer.7 Although tumor-associated carbohydrates (TACAs) are exploited as diagnostic markers,8 there is also evidence for naturally occurring antibodies to TACAs in healthy individuals.9
Insufficient responses to glycan-based vaccines or low titers of isohemagglutinins, antibodies to polysaccharide blood group antigens, are characteristic and diagnostic features of common variable immunodeficiency (CVID), the most frequent symptomatic antibody deficiency diagnosed in adulthood.10,11 Patients with specific antibody deficiency (SPAD) exhibit poor responses to structural or capsular polysaccharides of bacteria (eg, Streptococcus pneumoniae, Haemophilus influenzae), despite the presence of normal serum concentrations of immunoglobulin G (IgG), IgM, and IgA.12 Antibodies from different IgG subclasses are known to contain different specificities for glycan structures,9,13 which may explain the predisposition of certain patients with IgG subclass deficiency (IgGSD) to infections with encapsulated bacteria.12 However, current knowledge on the failure to raise adequate levels of glycan-specific antibodies in PAD is primarily based on experience with specific bacterial antigens.
The clinical assessment of PADs by diagnostic vaccination (eg, pneumococcal vaccines, Salmonella typhi Vi vaccine14 ) or the measurement of preexisting antibody titers (eg, isohemagglutinins) relies on a restricted number of glycan epitopes, thus providing only a narrow perspective of the actual degree of the immunodeficiency. Further disadvantages of diagnostic vaccination include diagnostic delay, interlaboratory variation, serotype-specific responses, age differences in antibody responses, or the challenging interpretation of preimmunization vs postimmunization specific antibody levels in patients not receiving IgG replacement therapy.10,11,15-18 The broader assessment of glycan-specific antibodies in patients may better reflect the immune defect and also facilitate treatment decisions, such as regarding life-long IgG-replacement therapy.
Glycan array technology allows the high-throughput analysis of specific antibody responses to carbohydrate antigens.19-21 In a previous study using glycan array version 5.1 of The Consortium for Functional Glycomics (CFG) to decipher the IgG repertoire of healthy individuals, we found that classes of glycans were recognized with different intensity, depending on the terminal carbohydrate moiety.9 Here, we used glycan array technology to investigate the IgG antibody repertoire of PAD patients in terms of clinically relevant carbohydrate epitopes, including microbial glycans, self-antigens, xenoantigens, and TACAs.
Materials and methods
Study design
This nonrandomized study was designed to investigate the human IgG anti-carbohydrate repertoire, in healthy and disease conditions, using glycan array technology combined with a computational system level approach. To this end, sera or purified IgG samples from healthy individuals or patients, as well as control antibodies, were screened on microarrays by the CFG or the US National Center for Functional Glycomics (NCFG).
Patient samples
Human blood was collected from healthy donors (HDs) or patients upon informed and written consent, in accordance with the Declaration of Helsinki. All experimental protocols were approved by the local institutional and/or licensing committees (KEK-BE: 148/10 and KEK No. 224/01). Therapeutic-naive patients followed at the University Hospital of Bern from January 2005 to December 2011 were retrospectively identified. Additional sera from therapeutic-naive patients without IgG-replacement therapy were provided by B.G. CVID was defined in accordance with the criteria of the Pan-American Group for Immunodeficiency and the European Society for Immunodeficiency.22 Inclusion criteria for IgGSD were a normal total IgG concentration with a significant decrease (>2 standard deviations below the mean for the age) in the serum concentrations of ≥1 IgG subclasses23 and recurrent episodes of infection. Symptomatic hypogammaglobulinemia (HGG) was defined as decreased total IgG concentration but not fulfilling the criteria for CVID with respect to a reduction in 2 Ig isotypes and/or reduced response to vaccination. For diagnostic vaccination, the pneumococcal polysaccharide vaccine (PPV) PNEUMOVAX 23 (MSD, Lucerne, Switzerland) was used. Levels of pneumococcal capsular polysaccharide (PCP) IgG > 50 mg/L and for PCP IgG2 > 40 mg/L were considered normal.
A sufficient PPV vaccination response was defined as a four-fold increase of postvaccination PCP IgG and PCP IgG2 titers and/or PCP IgG and IgG2 levels above 100 mg/L detected 4 to 6 weeks after vaccination. SPAD was diagnosed in patients with normal total Ig and IgG subclass concentrations but impaired PPV response. The characteristics of the different groups are summarized in supplemental Table 1 (available on the Blood Web site). Patients were not stratified for specific infections. The sera from these patients were individually screened in the NCFG glycan microarray version 2 (NCFGv2) or pooled in the indicated groups and screened in the CFG glycan array. IgG purification was performed by affinity chromatography in Ab SpinTrap columns (GE Healthcare). Pooled sera from patients and HDs were directly applied to the columns, and the purification procedures were performed according to the manufacturer’s instructions. The control IgG preparation (IgG control mix) consisted of 2 monoclonal human myeloma proteins, IgG1λ (67%) and IgG2κ (33%) (both from Sigma-Aldrich). This process resulted in a κ/λ ratio of 0.5, which is well within the range found in normal serum (0.26-1.65). The concentration of IgG in the different samples was determined using a Behring Turbitimer (Dade Behring). The quality of the isolated antibodies was checked by sodium dodecyl sulfate–polyacrylamide gel electrophoresis under reducing and nonreducing conditions (supplemental Figure 1).
Glycan array analysis
The glycan microarrays from the CFG (http://www.functionalglycomics.org/static/consortium/resources/resourcecoreh8.shtml) were prepared from amine functionalized glycan structures covalently coupled in microarrays to N-hydroxysuccinimide–derivatized microscope slides, as previously described.24 On the NCFGv2, carbohydrates were conjugated with the fluorescent linker AEAB, by reductive amination with sodium cyanoborohydrate, to form glycan-AEABs. The human sera or purified IgG or IgG control mix was processed identically for both arrays and screened at 180 μg/mL using a secondary biotinylated anti-human IgG monoclonal antibody (5 μg/mL; clone HP-6043), followed by Alexa Fluor 633–coupled streptavidin.9 A positive streptavidin control was used. Sample preparation and analysis were performed as indicated on the CFG Web site (www.functionalglycomics.org). To determine the specific binding to selected glycans, the antibody binding ratio (ABR) was calculated. The computed ABR represents the quotient of the respective sample relative fluorescent units (RFU) and the corresponding IgG control mix RFU. Data are expressed as the mean RFU or ABR values from 6 repeated experiments, unless indicated otherwise.
Database search
The identity or characteristics of glycans were investigated by consulting the CFG (http://www.functionalglycomics.org/fg/) or PubMed (http://www.ncbi.nlm.nih.gov/pubmed) database. The online Bacterial Carbohydrate Structure Database (BCSDB) was consulted to identify the bacterial origin of the glycans (http://csdb.glycoscience.ru/bacterial/).
Statistical analysis
Correlation matrixes, heat maps, and hierarchical clustering were performed using R (The R Foundation for Statistical Computing, version 3.0.2).25 Representation of the data was done using the heatmap.2 function in the package gplots.26 Hierarchical clustering and dendrograms were calculated with the complete method of the hclust function. Statistical analysis was performed using Microsoft Excel 2011 version 14.0.0 (Microsoft Corporation) or GraphPad Prism version 6.0c (GraphPad Software). For clique distribution analyses, only groups with a common terminal carbohydrate moiety and a minimal size of 12 glycans (2% of total) on CFG glycan array version 5.1 were considered. The Kruskal-Wallis test, Student t test, Spearman’s rank correlation, and 2-way analysis of variance were used for statistical analyses. Given the small size of the group (n = 2), the CVID with low PPV vaccination response (CVID PPVlow) subset on the NCFGv2 was excluded when group comparisons were performed.
Additional methods are available in supplemental Data.
Results
Broad carbohydrate reactivity defects in diverse PAD entities
Sera from PAD patients or HDs were screened on CFG glycan array version 5.1 to analyze the IgG-binding reactivity to 610 distinct glycans at 180 μg/mL. In accordance with previous studies,9,13 this assay concentration was determined to be optimal, resulting in reproducible IgG glycan-binding patterns with minimal background. Cohorts included patients with symptomatic HGG (n = 76), SPAD (n = 5), IgGSD (n = 8), CVID (n = 25), or CVID PPVlow (n = 6) and HDs (n = 43).
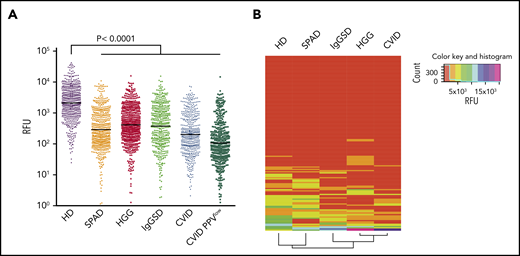
Using this platform, we observed broad IgG reactivity to printed glycans in pooled sera from HDs (Figure 1A; supplemental Table 2), which is consistent with previously published data.9,13,21,27-29 In contrast, a marked reduction in overall glycan-binding intensities was observed for all investigated PAD patient subsets, with the lowest averaged RFU in CVID, particularly in CVID PPVlow patients (Figure 1A). Next, IgG was purified from CVID, HGG, and HD sera by affinity chromatography and then screened on glycan array. As with whole sera, anti-glycan reactivity of equimolar purified IgG from immunocompromised patients exhibited lower mean RFU values compared with healthy controls (supplemental Figure 1).
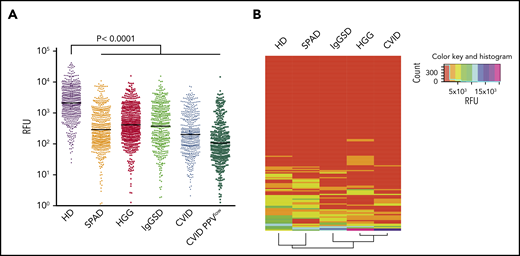
Antibody repertoire profiling on CFG and NCFGv2 reveals broad carbohydrate-recognition defects in symptomatic PADs. (A) Glycan-specific binding of serum IgG screened at 180 μg/mL on CFG glycan array version 5.1 (610 glycans) depicted as RFU. Screened sera were from cohorts with symptomatic HGG (n = 76), SPAD (n = 5), CVID (n = 25), CVID PPVlow (n = 6), or IgGSD (n = 8) or from HDs (n = 43). Significant values are reported (Kruskal-Wallis test). (B) Glycan-binding reactivity matrices for individual sera (HD, n = 12; SPAD, n = 5; IgGSD, n = 8; HGG, n = 11; CVID, n = 12) screened on the NCFGv2 and computed using the dendrogram clustering algorithm, as outlined in Materials and methods. Columns represent the antibody-reactivity profiles (reactivity of each specific glycan for the different sera samples), and the rows reflect the immune profiles for each patient subgroup based on the mean RFU values.
Antibody repertoire profiling on CFG and NCFGv2 reveals broad carbohydrate-recognition defects in symptomatic PADs. (A) Glycan-specific binding of serum IgG screened at 180 μg/mL on CFG glycan array version 5.1 (610 glycans) depicted as RFU. Screened sera were from cohorts with symptomatic HGG (n = 76), SPAD (n = 5), CVID (n = 25), CVID PPVlow (n = 6), or IgGSD (n = 8) or from HDs (n = 43). Significant values are reported (Kruskal-Wallis test). (B) Glycan-binding reactivity matrices for individual sera (HD, n = 12; SPAD, n = 5; IgGSD, n = 8; HGG, n = 11; CVID, n = 12) screened on the NCFGv2 and computed using the dendrogram clustering algorithm, as outlined in Materials and methods. Columns represent the antibody-reactivity profiles (reactivity of each specific glycan for the different sera samples), and the rows reflect the immune profiles for each patient subgroup based on the mean RFU values.
In an effort to compare the carbohydrate-specific IgG repertoires between PAD patients and healthy subjects, individual sera were screened. To this end, the NCFGv2 containing 147 glycans was used, which allowed us to include more samples. Identical glycans were similarly recognized on both arrays (supplemental Figure 2). Figure 1B illustrates the computed reactivity matrix (mean RFU values) ordered by a dendrogram clustering algorithm.30 Columns represent the antibody reactivity profiles (reactivity of each specific glycan for the different sera samples), and the rows reflect the immune profiles for each patient subgroup. The dendrogrammed reactivity matrix illustrates the broader spectrum of glycan reactivity in HD sera compared with all screened PAD patient subgroups, yet revealing a similar binding profile compared with the SPAD cohort. The glycan-binding profiles of IgGSD, HGG, and CVID sera displayed a narrower spectrum of glycan reactivity, which was most impaired in the CVID subgroup. CVID and CVID PPVlow patients also exhibited the lowest mean RFU values (supplemental Figure 2). No clear clustering of individual immunoprofiles was observed for total glycans, which was expected because the arrays contain synthetic glycans without known biological relevance (supplemental Figure 2).
Glycan-specific IgG antibodies to bacterial antigens in PAD
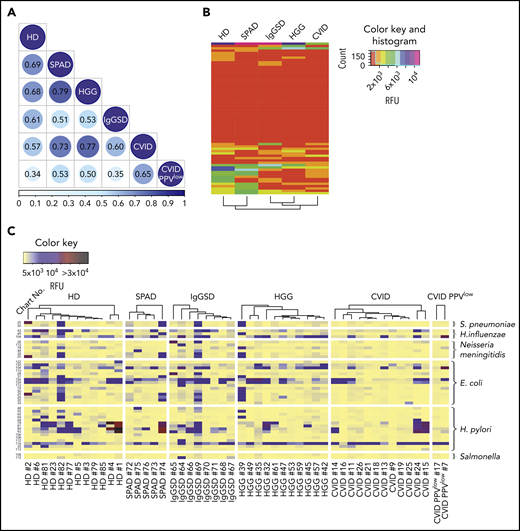
PADs are characterized by recurrent, severe, or unusual bacterial infections, especially by bacteria protected by a polysaccharide capsule. Thus, we decided to analyze the IgG reactivity profiles to known bacterial carbohydrates. On glycan array CFG version 5.1, we previously identified 121 bacterial carbohydrate antigens by consulting the BCSDB,9 providing a platform for high-throughput analysis for antibody reactivity to carbohydrate epitopes of multiple bacterial species. The printed glycans include capsular and structural oligosaccharides, as well as exopolysaccharides of commensal and pathogenic bacterial species.9 In comparison with immunoprofiles of healthy subjects, Spearman’s rank correlation analysis revealed divergent IgG reactivity profiles to bacterial glycan epitopes in PAD cohorts, with the lowest correlation for CVID PPVlow patients (0.34) and 0.57 for CVID, when pooled sera were screened on the CFG array (Figure 2A). Figure 2B illustrates the collective reactivity matrix (mean RFU values) of individual sera to 65 bacterial glycans on the NCFGv2. Hierarchical clustering analysis revealed a substantial loss of reactivity for the IgGSD, HGG and CVID cohorts. An extensive reduction in antibacterial reactivity was also found for recognition of the majority of 121 bacterial glycans on the CFG array version 5.1 in pooled sera from patients in these cohorts (supplemental Figure 3). No clear clustering of individual immunoprofiles was observed for bacterial glycans, which may be due to differential natural exposure to bacteria (supplemental Figure 3).
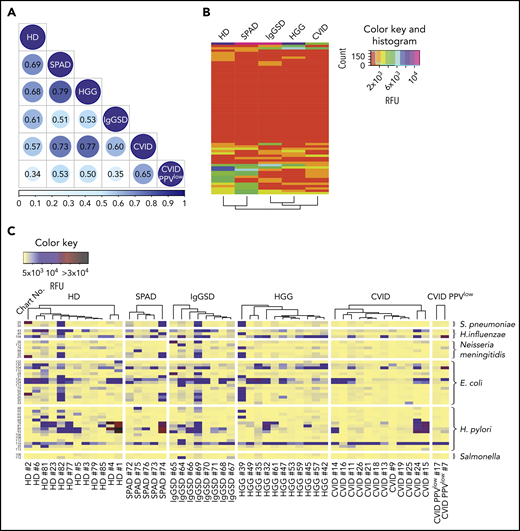
Recognition of bacterial carbohydrate epitopes in PADs. (A) Spearman’s rank correlation matrix for IgG recognition of pooled sera to all 121 bacterial carbohydrate structures identified by BCSDB analysis on the CFG array. (B-C) Recognition of bacterial glycans by individual sera screened on the NCFGv2. (B) Dendrogrammed reactivity matrix based on the mean RFU values for all 65 glycans identified by BCSDB analysis. (C) Heat map presentation of glycan-specific IgG binding to selected epitopes from S pneumoniae (n = 2), H influenzae (n = 3), N meningitidis (n = 6), E coli (n = 15), H pylori (n = 15), and Salmonella spp (n = 2).
Recognition of bacterial carbohydrate epitopes in PADs. (A) Spearman’s rank correlation matrix for IgG recognition of pooled sera to all 121 bacterial carbohydrate structures identified by BCSDB analysis on the CFG array. (B-C) Recognition of bacterial glycans by individual sera screened on the NCFGv2. (B) Dendrogrammed reactivity matrix based on the mean RFU values for all 65 glycans identified by BCSDB analysis. (C) Heat map presentation of glycan-specific IgG binding to selected epitopes from S pneumoniae (n = 2), H influenzae (n = 3), N meningitidis (n = 6), E coli (n = 15), H pylori (n = 15), and Salmonella spp (n = 2).
The glycan array technology, combined with consultation of the BCSDB databank, allowed us to screen in parallel several distinct epitopes that were identified as glycan constituents of specific bacteria. Figure 2C shows individual profiles of IgG reactivity to glycans on the NCFGv2 that are found in Escherichia coli (glycans: n = 15), Helicobacter pylori (n = 15), Neisseria meningitidis (n = 6), H influenzae (n = 3), S pneumoniae (n = 2), and Salmonella spp (n = 2). Heterogenous immunoprofiles were observed in all cohorts, including healthy individuals. However, reactivity profiles were particularly low in CVID and CVID PPVlow patients, who exhibited a significant loss of IgG antibodies specific for pathogens and commensals, including antigens to which human beings are frequently exposed and immunized. Missing antibodies against Neisseria meningitis and Salmonella in the different groups may reflect lack of exposure to these pathogens.
Recognition of TACA and Siglec ligands in PAD
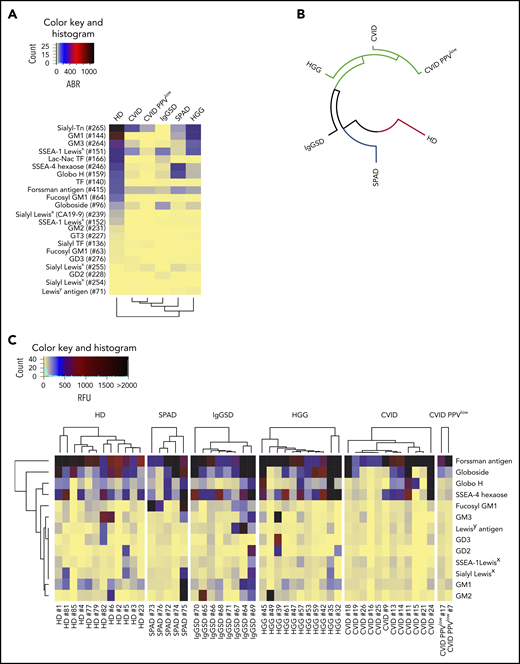
One life-threatening complication of PAD, in particular of CVID, is the occurrence of malignancies.1,31-34 Because altered surface glycosylation is a hallmark of cancer and influences different aspects of tumor progression and antitumor immunity,7,35,36 we set out to explore IgG reactivity to tumor-associated carbohydrate antigens. Figure 3A demonstrates the IgG reactivity profiles to 22 glycan epitopes on CFG glycan array version 5.1 that constitute well-established TACAs. In accordance with our previous data,9 HD sera contained natural antibodies against TACAs. High reactivity was found against Lewis antigen–related TACAs, gangliosides, Thomsen-Friedenreich antigen, and globo-series–associated antigens. Loss of anti-TACA IgG reactivity was found in all examined PAD patient subgroups. The reduced binding activity by IgG was most dominant in CVID PPVlow donors, followed by CVID patients, as computed based on RFU profiles and represented as a circular dendrogram (Figure 3B). To further explore these findings, we analyzed anti-TACA signatures of individual sera screened on the NCFGv2. Most sera from healthy individuals exhibited high anti–TACA reactivity to Forssman antigen, globoside, Globo H, and SSEA-4 hexaose but heterogenous binding to other TACAs (Figure 3C). The substantial loss of anti-TACA reactivity in CVID was confirmed in individual sera. In contrast, antibody binding to the Forssman antigen was found in all cohorts and all screened individual sera (Figure 3A,C), indicating that this glycan epitope is highly immunogenic with widely maintained specific antibody production in PAD patients. Indeed, in CVID PPVlow patients and in some CVID patients, Forssman antigen was the only investigated TACA for which higher-intensity signals were detected. Reactivity to GM1 was particularly low in CVID patients. This is remarkable because GM1 expression is present and positively correlates with rituximab treatment response in non-Hodgkin lymphoma,37 which is among the most frequent malignancies occurring in these patients.10
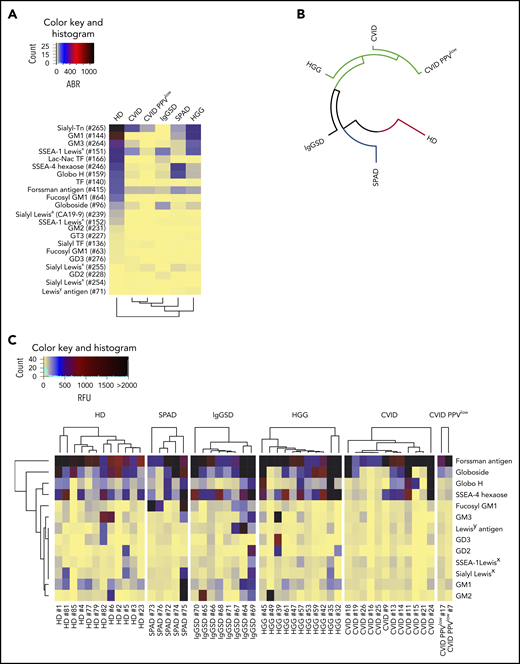
Recognition of TACAs by glycan-specific IgG in PAD. Recognition of TACAs by pooled serum IgG, as screened on the CFG array and depicted as dendrogrammed glycan reactivity matrix based on ABR (A) or as a radial dendrogram based on RFU values (B). (C) Heat map presentation based on RFU values illustrating the reactivity of individual serum IgG to TACAs, as revealed by screening on the NCFGv2.
Recognition of TACAs by glycan-specific IgG in PAD. Recognition of TACAs by pooled serum IgG, as screened on the CFG array and depicted as dendrogrammed glycan reactivity matrix based on ABR (A) or as a radial dendrogram based on RFU values (B). (C) Heat map presentation based on RFU values illustrating the reactivity of individual serum IgG to TACAs, as revealed by screening on the NCFGv2.
Overexpression of sialylated TACAs frequently occurs in tumors, and hypersialylation has been linked to tumor progression and immune escape by the engagement of inhibitory Siglec receptors on leukocytes.7,36,38-40 In PAD cohorts, the antibody recognition of certain sialoglycans with known Siglec-binding capacity was similar or reduced on the glycan array compared with HD responses (supplemental Figure 4). However, for 2 ligands of CD22/Siglec-2 expressed on B cells, binding was high (glycan #377) or even increased (glycan #268). Little is known about the tissue expression of these 2 ligands, and it remains to be shown whether such antibodies at higher levels might interfere with the CD22 receptor/ligand axis to unleash B-cell responses under certain circumstances.
Failure to raise Galα- and GalNAc-directed IgG antibodies in PAD
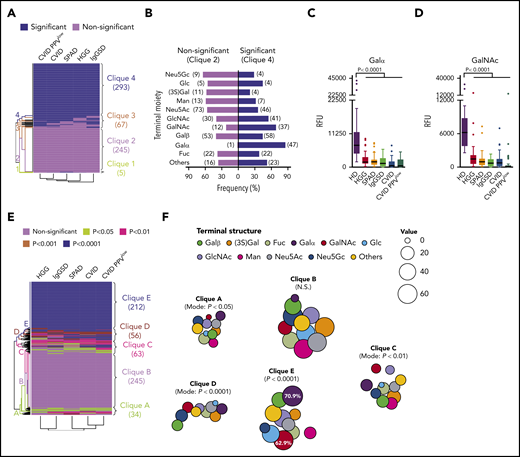
The composition of glycan antigens, in particular the terminal carbohydrate moiety, has been linked to the immunogenicity of a given glycan in healthy individuals.9 This raised the question about the effect of the relationship between immune system failure (ISF) in PAD subgroups and the structure-related immunogenicity on the architecture of the glycan-specific IgG repertoire. A binary deviation matrix was computed considering statistical deviation (P < .05) of IgG binding (RFU values) for each PAD cohort and each glycan compared with HD data and reordered by a dendrogram clustering algorithm (Figure 4A). The rows in this matrix indicate the binary antibody-reactivity profiles, and the columns represent the deviation immune profiles for each PAD subgroup. By hierarchical clustering analysis 4 major subgroups were identified: predominant were clique 4 representing glycans with deviation in all PAD cohorts (n = 293; 48%) and clique 2 containing glycans without deviation in all PAD cohorts (n = 245; 40%); of lower magnitude were clique 1 (n = 5; 1%) and clique 3 (n = 67; 11%) encompassing glycans with heterogeneous IgG-binding activities between subgroups (supplemental Table 3).
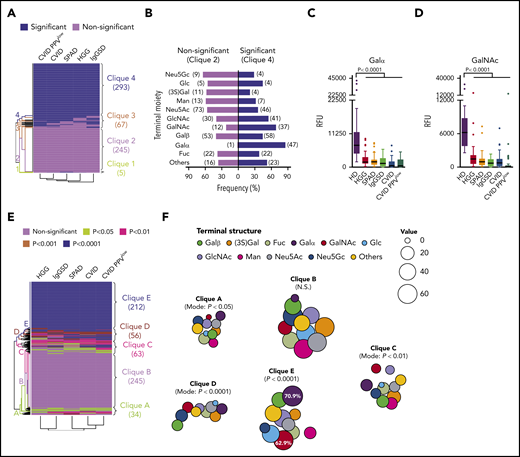
Deviant recognition of distinct terminal carbohydrate moieties and specific loss of Galα reactivity in symptomatic PAD. (A) Binary deviation map (P < .05) of IgG immunoprofiles compared with healthy controls based on 2-way analysis of variance and hierarchical clustering analysis with identification of cliques 1 to 4. The rows in this matrix represent the binary antibody-reactivity profiles, and the columns represent the deviation immune profiles for each PAD subgroup. (B) Terminal carbohydrate moieties of epitopes in reactivity clique 2 (nonsignificant deviation) and clique 4 (significant deviation). Numeric occurrence is indicated in parenthesis. Recognition of Galα (C) or GalNAc (D) epitopes within clique 4 (significant deviation). (E) Deviation map based on degree of significance with identification of cliques A through E. (F) Bubble chart displaying terminal glycan structure distribution across cliques A through E. The size key and percentage numbers represent the frequencies.
Deviant recognition of distinct terminal carbohydrate moieties and specific loss of Galα reactivity in symptomatic PAD. (A) Binary deviation map (P < .05) of IgG immunoprofiles compared with healthy controls based on 2-way analysis of variance and hierarchical clustering analysis with identification of cliques 1 to 4. The rows in this matrix represent the binary antibody-reactivity profiles, and the columns represent the deviation immune profiles for each PAD subgroup. (B) Terminal carbohydrate moieties of epitopes in reactivity clique 2 (nonsignificant deviation) and clique 4 (significant deviation). Numeric occurrence is indicated in parenthesis. Recognition of Galα (C) or GalNAc (D) epitopes within clique 4 (significant deviation). (E) Deviation map based on degree of significance with identification of cliques A through E. (F) Bubble chart displaying terminal glycan structure distribution across cliques A through E. The size key and percentage numbers represent the frequencies.
Given the association between immunogenicity and terminal carbohydrate moiety of glycans,9,41 the clique distribution of glycans was investigated based on their structure. The most dominant deviation was found for Galα- and GalNAc-terminated glycans (Figure 4B). Forty-seven (85.5%) vs only 1 (1.8%) Galα structure(s) and 37 (68.5%) vs 12 (22.2%) GalNAc-terminated glycans were represented in clique 4 and clique 2, respectively. In accordance, the IgG antibody-binding levels (RFU values) of Galα- and GalNAc-terminated glycans were lower in all cohorts (Figure 4C-D). In-depth analysis computing a deviation matrix based on P values and hierarchical clustering analysis revealed large glycan clusters that were concomitantly nonaberrant (clique B) or aberrant (clique E) for all disease entities compared with healthy individuals (Figure 4E), whereas Galα-terminated (70.9%) and GalNAc-terminated (62.9%) glycans were most prevalent in clique E (Figure 4F), indicating failed antibody responses to these specific structures.
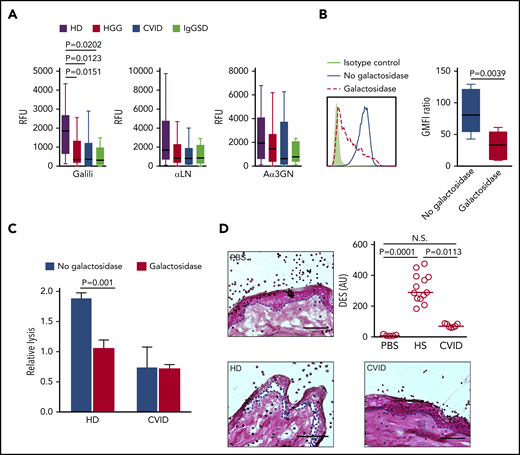
As a consequence of the inactivation of the GGTA1 gene encoding for α1,3galactosyltransferase, humans, apes, and Old World monkeys do not express the Galili epitope Galα1-3Galβ1-4GlcNAcβ.42 Natural antibodies to this xenoantigen in most humans, eventually generated in response to the microbiota of the host, are considered a key factor in the rejection of xenografts43 and necessitated the development of α1,3galactosyltransferase-knockout animals.44,45 Using a suspension array, we examined individual sera from patients (HGG, n = 37; CVID, n = 15; IgGSD, n = 8) and HDs (n = 18) for IgG reactivity to the Galili epitope and other Galα- and GalNAc-terminated structures, including αLN (Galα1-4GlcNAcβ), Aα3GN (Galα1-3GlcNAcβ), or A and B blood group antigens (Figure 5A; supplemental Figure 5). Low isohemagglutinins are frequently observed in CVID patients.11 Consistent with this observation, we also found reduced IgG reactivity to blood group A and B antigens in the HGG, SPAD, and IgGSD cohorts (supplemental Figure 6). IgG reactivity to Galα- and GalNAc-terminated epitopes was consistently lower in most PAD patients; the reduction was most significant for the Galili epitope, as evidenced by the statistical analysis (Figure 5A; supplemental Figure 5).
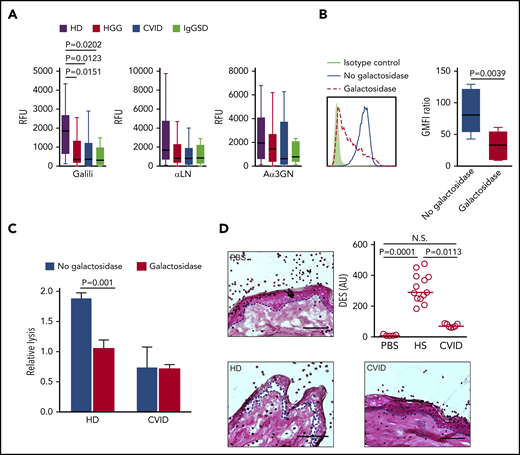
Diminished recognition of Galα-terminated glycan epitopes and reduced Galα-dependent xenogeneic anti-porcine reactivity of CVID and symptomatic IgGSD sera. (A) IgG antibody reactivity to Galili, αLN, and Aα3GN glycans, as assessed by suspension array (multiplex immunoassay). Individual sera from HDs (n = 18) or from HGG (n = 37), CVID (n = 15), and IgGSD (n = 8) patients were analyzed. Box-and-whisker diagrams were created by the Tukey method. The middle line is plotted at the median, and the box represents the interquartile range (difference between the 25th and 75th percentiles). Significant values are reported (Kruskal-Wallis test). (B) Reduced surface staining of anti-Galili monoclonal antibody reactivity following α-galactosidase treatment of porcine PK15 cells, as assessed by flow cytometry. Representative line graph (left panel) and summary (right panel). Student t test was used to calculate the P values. (C) ADCC activity of primary human NK cells against PK15 cells at an effector-to-target ratio of 10:1, in the absence or presence of α-galactosidase (n ≥ 3). Significant values are reported (Student t test). (D) Histopathologic analysis of pig skin damage in cryosections incubated with healthy and patient sera in the presence of healthy human leukocytes. Representative examples for each analyzed disease and control are shown (left panels and lower right panel; hematoxylin and eosin stain). The dashed lines indicate the dermal–epidermal junction. Scale bars, 75 µm. The damage induced was calculated as the dermal–epidermal separation (DES) normalized to the IgG concentration in the sera (upper right panel) (n ≥ 7). Significant values are reported (Kruskal-Wallis test). N.S., not significant; PBS, phosphate-buffered saline.
Diminished recognition of Galα-terminated glycan epitopes and reduced Galα-dependent xenogeneic anti-porcine reactivity of CVID and symptomatic IgGSD sera. (A) IgG antibody reactivity to Galili, αLN, and Aα3GN glycans, as assessed by suspension array (multiplex immunoassay). Individual sera from HDs (n = 18) or from HGG (n = 37), CVID (n = 15), and IgGSD (n = 8) patients were analyzed. Box-and-whisker diagrams were created by the Tukey method. The middle line is plotted at the median, and the box represents the interquartile range (difference between the 25th and 75th percentiles). Significant values are reported (Kruskal-Wallis test). (B) Reduced surface staining of anti-Galili monoclonal antibody reactivity following α-galactosidase treatment of porcine PK15 cells, as assessed by flow cytometry. Representative line graph (left panel) and summary (right panel). Student t test was used to calculate the P values. (C) ADCC activity of primary human NK cells against PK15 cells at an effector-to-target ratio of 10:1, in the absence or presence of α-galactosidase (n ≥ 3). Significant values are reported (Student t test). (D) Histopathologic analysis of pig skin damage in cryosections incubated with healthy and patient sera in the presence of healthy human leukocytes. Representative examples for each analyzed disease and control are shown (left panels and lower right panel; hematoxylin and eosin stain). The dashed lines indicate the dermal–epidermal junction. Scale bars, 75 µm. The damage induced was calculated as the dermal–epidermal separation (DES) normalized to the IgG concentration in the sera (upper right panel) (n ≥ 7). Significant values are reported (Kruskal-Wallis test). N.S., not significant; PBS, phosphate-buffered saline.
Given the significant loss of anti-Galili reactivity in PAD patients, functional implications were tested in a xenoreactivity assay, analyzing antibody-dependent cell cytotoxicity (ADCC) of primary human natural killer (NK) cells directed against the porcine kidney cell line PK15. The Galili epitope is highly expressed on the surface of this cell line and is lost following enzymatic digestion by α-galactosidase, as assessed by flow cytometry (Figure 5B). Sera from HDs induced substantial NK cell–mediated ADCC activity against the Galili+ PK15 cells, which was abolished following α-galactosidase treatment of the porcine target cells (Figure 5C); this illustrated the dependence of the xenogeneic activity on Galα epitopes, including Galili. In contrast, CVID sera failed to promote NK cell–mediated ADCC against the porcine Galili+ target cells at equimolar IgG concentrations (1 mg/mL), indicating a repertoire defect in the recognition of xenogeneic antigens. Moreover, loss of the xenogeneic potential of CVID sera was demonstrated in a modified version of an established model for antibody-mediated skin damage,46,47 involving cryosections of porcine skin incubated with patients’ sera and leukocytes from HDs and the assessment of dermal–epidermal separation. Porcine skin damage was significantly lower in the presence of CVID sera compared with human control sera at equimolar IgG concentrations (Figure 5D). Together, these data provide evidence for humoral immune system failure beyond quantitative antibody deficiency, which involves repertoire defects in PAD.
Discussion
The emerging picture is that the human IgG anti-carbohydrate repertoires in health,9 as well as in primary antibody deficiencies, exhibit a modular organization, yet with a different architecture in PAD. Translated to Cohen’s concept of immune computation,48 our data imply that primary antibody deficiency is not solely characterized by aberrant antibody production; rather, it features disturbed immune-system response states (the output) as the result of an altered algorithm to compute immunogenic states (the input) that is dictated by impaired cellular and molecular networks of innate and adaptive immunity.49,50
Despite the heterogeneous genetic basis across PAD entities,1 deviation mapping revealed significant repertoire similarities pointing toward a common disease-modifying algorithm that drives humoral immune system failure. Dominant was the reduced or lost specificity for Galα- or GalNAc-terminated glycan epitopes in PAD patients, to which healthy individuals express high levels of naturally occurring antibodies, eventually in response to immune stimulation by carbohydrate antigens of their microbiota.9,41 Thus, the loss of Galα- or GalNAc-specific antibodies might reflect the lost capacity to induce these antibodies in response to the host microbiota, the microbial dysbiosis in PAD,4 or both. Interestingly, bacteria and tumor cells use certain glycosylation patterns to escape immunity, such as sialoglycans that engage inhibitory Siglec receptors on leukocyte subsets of myeloid and lymphoid lineages.6,35,39,40,51 It remains to be shown whether naturally occurring antibodies to TACA are also induced by the microbiota, as well as whether the observed loss has consequences for PAD patients. Notably, the loss of IgG reactivity to tumor antigens was most evident in the CVID cohort, a disorder in which the incidence of malignancy is increased,10 supporting the concept of aberrant immune surveillance in primary immunodeficiency.52 For instance, we observed low antibody reactivity to TACAs known to be expressed on CVID-associated malignancies, such as GM1 in non-Hodgkin lymphoma37 or Thomson-Friedenreich antigen in gastric cancer.53,54 However, tumor glycosylation patterns and their immunoreactivity remain to be fully explored for CVID-associated malignancies.
Glycoimmunology remains a relatively underdeveloped field, and the need for interdisciplinary collaborations and enhanced training of glycoscientists in biomedical areas has been identified.55 In line with this, there is also a demand for tools, such as glycan arrays; their development is challenging given the availability of differently synthesized or purified glycans. However, this initial study highlights the potential and the need to expand glycan array studies in PAD using biological samples from larger patient cohorts. Clusters of glycans were identified with different reactivity profiles between PAD subgroups (eg, clique 3 or cliques A, C, D), which may be candidates for diagnostic use. In future studies, it will also be advisable to compare glycan-specific antibody profiles with clinical and laboratory parameters (eg, switched memory B-cell numbers, blood groups) and in specifically classified patient subsets.56
The broad and qualitative assessment of ISF in individual patients may be informative with regard to the severity of the disease, eventually leading to more personalized treatment decisions. Thus, glycan array technology may provide a useful option for the individual assessment of ISF and facilitate clinical practice, especially given the heterogeneity of PAD entities with a growing list of identified pathogenetic defects.2 However, our broad analysis demonstrates the extent of the skewed IgG repertoire with impaired reactivity to biologically relevant glycan epitopes, including those linked to PAD-associated clinical manifestations, such as infection, malignancy, and autoimmunity.1,10 Our systems immunology approach highlights the power of high-throughput assessment of humoral immune system failure by microarray technology, which may have potential ramifications for the diagnosis, classification, and therapy of PAD patients.
For original data, please contact Stephan von Gunten (stephan.vongunten@pki.unibe.ch).
The online version of this article contains a data supplement.
The publication costs of this article were defrayed in part by page charge payment. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734.
Acknowledgments
The authors thank Tatiana Pochechueva (Gynecology, University Hospital, Basel, Switzerland) for assistance with the suspension array, Alexander Chinarev (Shemyakin-Ovchinnikov Institute of Bioorganic Chemistry, Russian Academy of Sciences, Moscow) for monobiotinylated glycopolymer synthesis. The authors also gratefully acknowledge Alyssa McQuillan, Akul Mehta, and Jamie Heimburg-Molinaro (Department of Surgery, Beth Israel Deaconess Medical Center, Harvard Medical School, Boston, MA) for technical assistance with printing the glycan microarrays and conducting assays and analyses. The authors thank Yuniel Fernandez-Marrero (Institute of Pharmacology, University of Bern, Bern, Switzerland) for assistance with statistical analyses.
This work was supported in part by the Swiss National Science Foundation (grants 310030_162552 and 310030_184757) (S.v.G.), Swiss Cancer League/Swiss Cancer Research grants KFS-3941-08-2016 and KFS-3248-08-2013 (S.v.G.), and National Institutes of Health/National Institute of General Medical Sciences grants P41GM103694 and GM098791 (R.D.C.). H.-U.S. received support from the Swiss National Science Foundation (grants 310030-166473 and 310030_184816) and the European Union's Horizon 2020 research and innovation program (Marie Sklodowska-Curie grant 642295; MEL-PLEX). Work by N.B. was supported by Russian Scientific Foundation grant 14-50-00131. C.J. received support from the Swiss National Science Foundation (grants PMPDP3_129022 and PZ00P3_161459).
Authorship
Contribution: S.v.G. and P.J. designed the study; K.F.B., P.J., and S.v.G. wrote the manuscript; K.F.B. and S.v.G. analyzed the data; D.F.S. and R.D.C. supervised glycan array experiments conducted at the CFG; K.F.B. performed database searches and computational analysis of the data set; B.G., C.J., and P.J. collected, classified, and preparated patient samples; K.F.B., E.d.G., M.M.A., and A.D. performed experiments under the supervision of D.S., H.-U.S., R.R., and S.v.G; N.B. supervised glycan synthesis; and all authors had full access to the data, helped to draft or to critically revise the manuscript, contributed to data interpretation, and reviewed and approved the final version of the manuscript.
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Correspondence: Stephan von Gunten, Institute of Pharmacology, University of Bern, Inselspital INO-F, CH-3010 Bern, Switzerland; e-mail: stephan.vongunten@pki.unibe.ch.