In this issue of Blood, report that ARIEL (ARID5B-inducing enhancer-associated long noncoding RNA) is a novel enhancer RNA (eRNA) that amplifies oncogenic transcriptional loops in a subset of T-cell acute lymphoblastic leukemia (T-ALL).1
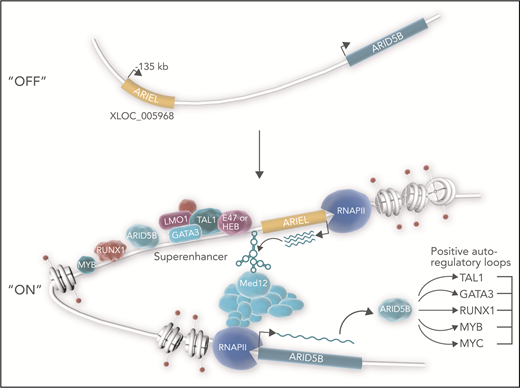
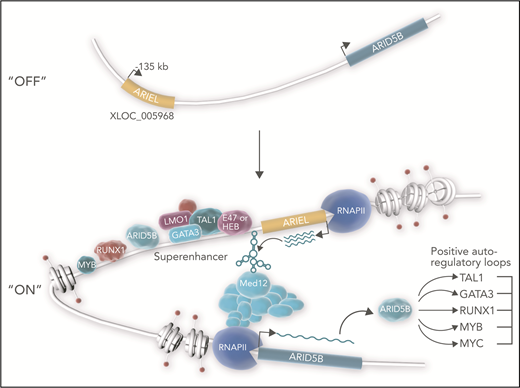
ARIEL (ARID5B-inducing enhancer-associated long noncoding RNA/XLOC_005968) acts as an eRNA in a subset of TAL1-type T-ALL. The TAL1 complex in T-ALL consists of several transcriptional regulators that include the class II basic helix-loop-helix transcription factor TAL1, E proteins (eg, E47 or HEB), LIM-only proteins (eg, LMO1/2), and GATA factors (eg, GATA3). This complex frequently co-occupies enhancers with partner transcription factors ARID5B, RUNX1, and MYB. In the nucleus, the TAL1 complex instructs RNA polymerase II (RNAPII) to transcribe ARIEL from the ARID5B enhancer template located at −135 kb relative to the ARID5B transcriptional start site.1,6 ARIEL connects the enhancer to MED12, a subunit of the mediator coactivator complex that binds RNAPII. Thus, ARIEL facilitates chromatin looping interactions between the enhancer and the ARID5B promoter. These interactions in turn promote the expression of ARID5B, which amplifies positive autoregulatory loops that generate the TAL1 oncogenic transcriptional program.7,8 Upregulated expression of ARID5B target genes promotes ARIEL transcription. In this diagram, direct interactions involving ARIEL and genomic coordinates of factors are postulated.
ARIEL (ARID5B-inducing enhancer-associated long noncoding RNA/XLOC_005968) acts as an eRNA in a subset of TAL1-type T-ALL. The TAL1 complex in T-ALL consists of several transcriptional regulators that include the class II basic helix-loop-helix transcription factor TAL1, E proteins (eg, E47 or HEB), LIM-only proteins (eg, LMO1/2), and GATA factors (eg, GATA3). This complex frequently co-occupies enhancers with partner transcription factors ARID5B, RUNX1, and MYB. In the nucleus, the TAL1 complex instructs RNA polymerase II (RNAPII) to transcribe ARIEL from the ARID5B enhancer template located at −135 kb relative to the ARID5B transcriptional start site.1,6 ARIEL connects the enhancer to MED12, a subunit of the mediator coactivator complex that binds RNAPII. Thus, ARIEL facilitates chromatin looping interactions between the enhancer and the ARID5B promoter. These interactions in turn promote the expression of ARID5B, which amplifies positive autoregulatory loops that generate the TAL1 oncogenic transcriptional program.7,8 Upregulated expression of ARID5B target genes promotes ARIEL transcription. In this diagram, direct interactions involving ARIEL and genomic coordinates of factors are postulated.
As students, many of us learned from the central dogma of molecular biology that information flows from DNA to RNA to protein. In the modern era, genome-scale sequencing has revealed that more than 70% of the genome is transcribed into RNAs that do not produce protein. These RNAs are called noncoding RNAs (ncRNAs). eRNAs have many other names and ambiguous definitions but can be considered simply as ncRNAs that are transcribed from enhancer elements.2 eRNAs were initially identified through transcriptome sequencing studies, which estimate their numbers in the ∼40 000 to 65 000 range. However, conventional sequencing underestimates the number of eRNAs because they are rapidly degraded by the RNA exosome. In contrast, specialized methods such as BruUV-seq in which UV light suppresses RNA exosome activity are more sensitive in detecting eRNAs.3 Many eRNAs lack function, but others can activate neighboring genes through cis-mediated mechanisms to regulate diverse cellular processes.
In 2014, eRNAs captured the attention of researchers in the field of T-ALL in a report that described the first comprehensive list of long ncRNAs and the first functional eRNA (LUNAR1) in human T-ALL.4 Subsequent studies confirmed the large-scale existence of eRNAs in T-ALL.5,6 But after 2014, no functionally important eRNAs were identified. Doubts grew. Perhaps LUNAR1 was a one-hit wonder. Perhaps eRNAs were simply transcriptional byproducts of enhancer activation. These doubts are now gone. Using a stunning variety of sophisticated techniques, Tan et al identified the eRNA XLOC_005968, which they named “ARIEL,” as a new major player in the TAL1-activated subset of T-ALL.
Tan et al previously reported that the transcription factor ARID5B was an important downstream target of the oncogenic TAL1 complex.7 TAL1 is aberrantly overexpressed in more than 40% of human patients with T-ALL. The TAL1 complex consists of several transcriptional regulators that co-occupy enhancers with partner transcription factors to activate positive feed-forward loops that induce oncogenes such as MYB and MYC.7,8 We learned that the TAL1 complex was the key that turned the circuit on. However, there were missing pieces. How does the TAL1 complex connect enhancers to promoters in three-dimensional space and engage the transcriptional machinery? Tan et al show that ARIEL seems to be a missing piece.
Tan et al report that the TAL1 complex and partners bind an enhancer at −135 kb from the transcriptional start site of ARID5B (see figure), which induces transcription of ARIEL from the enhancer element.6 Accordingly, ARIEL has higher expression in TAL1-type T-ALL cells than other T-ALL cells. Several carefully performed assays (eg, 4C-seq, chromatin immunoprecipitation polymerase chain reaction [PCR], chromatin isolation by RNA purification PCR, RNA immunoprecipitation, and RNA pulldown) showed that ARIEL binds mediator (a transcriptional coactivator complex), the ARID5B promoter, and the −135-kb enhancer. To test whether ARIEL is essential for assembling the ARID5B enhancer-promoter structure, Tan et al knocked ARIEL down. Loss of ARIEL reduced enhancer-promoter interactions and enhancer occupancy of TAL1 complex members, partner proteins, mediator proteins, RNA polymerase II, and ARID5B itself. Furthermore, ARIEL knockdown reduced the expression of ARID5B and TAL1 target genes and impaired leukemic proliferation in vitro and in vivo. In other words, take ARIEL away and large protein-DNA complexes fall apart, oncogenic transcriptional loops unravel, and leukemic cells die.
Importantly, the studies by Tan et al were rigorous. The use of multiple orthogonal methods raises confidence in the proposed dynamic structural effects of ARIEL on the ARID5B locus (see figure). Rigor was particularly evident when the authors showed that ARIEL had strong effects on some specific proteins and genes (but not others) and in some TAL1-positive cell lines (but not others). The discoveries of LUNAR1 and ARIEL argue for a wider search for eRNAs that control T-ALL leukemogenesis. These elegant studies can serve as a basic framework for future experimental designs. However, there were limitations that should be noted. For example, despite numerous attempts, Tan et al were unable to test the potential of ARIEL as a therapeutic target through genetic silencing in primary patient-derived xenografts. These technical issues highlight a need to improve current protocols for widespread applicability.9,10 Another limitation is that it remains unclear which effects of ARIEL are direct. Mapping the direct eRNA-to-protein or eRNA-to-DNA interactions is challenging but would be useful for designing potential inhibitors that disrupt eRNA functions.
Tan et al discovered a key mechanism and vulnerability that amplifies the TAL1-driven oncogenic genetic program. However, the clinical implications remain speculative. There is currently no strategy to effectively target eRNAs. Synthetic nucleotides such as antisense oligonucleotides and small interfering RNAs could be designed to silence ARIEL expression. However, for decades, use of synthetic nucleotides to target RNAs has met formidable obstacles. In particular, in vivo delivery of synthetic nucleotides into cancer cells has been challenging. Furthermore, the potential toxicity of ARIEL inhibition is unknown, but it could be addressed using genetically modified animal models. Finally, the potential of ARIEL as a biomarker remains to be explored. It is unknown whether its expression correlates with patient outcomes. Because the explosive growth of new reports elucidating eRNA biochemistry is expected to continue in the near future, it is possible that new strategies for clinical translation will soon be revealed.
Mechanistically, ARIEL (as well as other eRNAs) remains enigmatic. For example, ARIEL acts as a cis regulator of ARID5B but also seems to have yet-to-be-revealed ARID5B-independent effects. In addition, yet-to-be-revealed context-dependent factors seem to regulate ARIEL expression because many TAL1+ T-ALLs do not express ARIEL. Finally, candidate approaches that test individual eRNAs have met with some success but have not generated the momentum needed to radically advance the field. It will be important to devise large-scale functional genomic studies to screen for oncogenic eRNAs. The ARID5B-activated circuit is only 1 part of a vast sea of circuits. Tan et al found ARIEL, but there are more leukemia-associated eRNAs to discover. For now, we eagerly wait to learn what other eRNAs lie under the sea of circuits that support T-ALL.
Conflict-of-interest disclosure: The author declares no competing financial interests.