

In this paper, 1 analyzed Burkitt lymphoma cells by using a relatively new, general method for finding potential therapeutic target genes in cancer. They showed that the protein product of the CYB561A3 gene is the key lysosomal iron reductase that is necessary for Burkitt B-cell growth and survival. A specific inhibitor of the CYB561A3 protein may therefore be a candidate gene for Burkitt lymphoma therapy.
The experimental approach involved lentivirus expression of libraries of CRISPR single-guide RNAs (sgRNAs) in a Burkitt lymphoma cell line and in a suitable control cell line. An Epstein-Barr virus (EBV)-transformed B lymphoblastoid cell line was used as the control. After the resulting populations of cells grew out, sequencing revealed which guide RNAs were depleted specifically in the Burkitt lymphoma cell line; therefore, the genes they targeted were necessary for growth or survival of those cells. The sgRNA libraries contain multiple sgRNAs per messenger RNA, so finding that all 4 CYB561A3 sgRNAs were missing gave a good degree of confidence that this gene would be essential in the Burkitt lymphoma cells. There is already a database of such screenings in many different cancer cell lines (DepMap2) and this supported the uniqueness of the role of CYB561A3 in the several Burkitt lymphoma cell lines (with or without EBV) that were tested. Three other lymphoma cell lines in DepMap were also dependent on CYB561A3, but to a lesser degree.
Knockout of CYB561A3 in the Burkitt lymphoma cells caused a G2M arrest of the cell cycle and lysosome and mitochondrial damage, with eventual cell death. RNA sequencing analysis of Burkitt cells after knockdown of CYB561A3 showed strong induction of the transferrin receptor TFRC, indicating a loss of bioavailable iron in the cell. Correct iron metabolism with maintenance of a suitable balance of Fe2+ and Fe3+ is known to be essential for cell survival.3 Detailed analysis showed that the key difference between Burkitt lymphoma cells and the control lymphoblastoid cell lines is in the 2 different pathways they use to maintain iron reduction levels. In lymphoblastoid cell lines, the STEAP3 protein uses reduced NAD phosphate as an electron donor for this, but the Burkitt lymphoma cells depend on CYB561A3, which uses ascorbate as its electron donor (see figure).
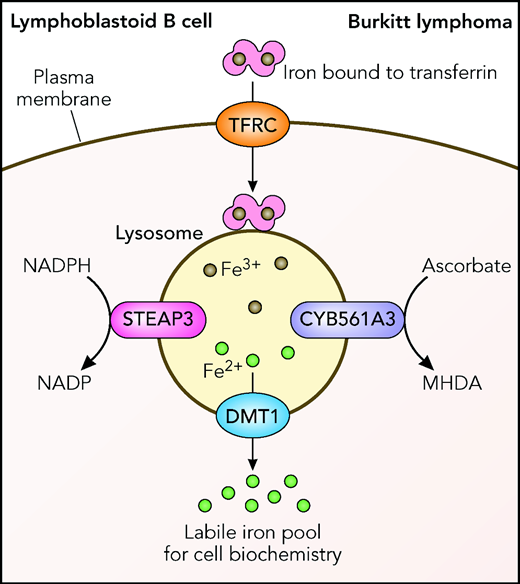
Transport of Fe3+ iron into cells on transferrin via the TFRC transferrin receptor and reduction of Fe3+ in the lysosome to Fe2+, either by STEAP3 or by CYB561A3. The divalent metal transporter DMT1 transfers Fe2+ into the cytoplasm to participate in further cell metabolism. The figure has been adapted from Figure 7 in the article by Wang et al, which begins on page 2216. Professional illustration by Patrick Lane, ScEYEnce Studios.
Transport of Fe3+ iron into cells on transferrin via the TFRC transferrin receptor and reduction of Fe3+ in the lysosome to Fe2+, either by STEAP3 or by CYB561A3. The divalent metal transporter DMT1 transfers Fe2+ into the cytoplasm to participate in further cell metabolism. The figure has been adapted from Figure 7 in the article by Wang et al, which begins on page 2216. Professional illustration by Patrick Lane, ScEYEnce Studios.
MYC is a transcription factor, whose gene is characteristically subject to a chromosomal translocation in Burkitt lymphoma,4 with consequent deregulated expression of MYC protein. In addition to its effects on cell proliferation and apoptosis, MYC regulates several genes involved in iron metabolism, including TFRC and the ferrous iron transporter DMT1.5 There is therefore an inherent link between Burkitt oncogenesis and iron metabolism that may be manifested in the apparently unique dependence of Burkitt cells on the CYB561A3 pathway. Iron deficiency is frequent in patients who have holoendemic malaria,6 which is a key risk factor for endemic Burkitt lymphoma. Wang et al therefore speculate that CYB561A3 ferrireductase activity offers a selective advantage to EBV-infected B cells at a particular stage of transformation, perhaps also relevant to development of endemic Burkitt lymphoma.
There is currently no specific inhibitor of the CYB561A3 iron reductase available and further validation of the unique role of CYB561A3 will be needed, for example, in knockout mice before preclinical development. There is a long way to go before creating a CYB561A3 targeted drug for Burkitt lymphoma, but Wang et al offer an excellent demonstration of the power of the sgRNA library approach to identify cell proteins that could become specific targets for novel cancer therapies.
Conflict-of-interest disclosure: The author declares no competing financial interests.

