

Abstract
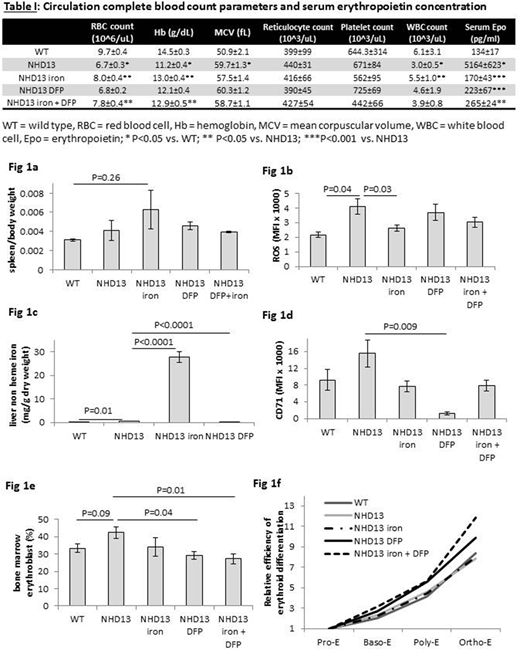
Myelodysplastic syndrome (MDS) is a heterogeneous group of bone marrow stem cell disorders characterized by ineffective hematopoiesis, cytopenias, and transformation to acute leukemia (AML). Low risk MDS patients exhibit a longer median survival, the lowest rate of progression to AML, and account for approximately two-thirds of all MDS patients, requiring recurrent RBC transfusions to alleviate symptomatic anemia. Transfusion-dependence results in iron overload which is associated with reduced overall survival. Methods to diagnose and treat (e.g. iron chelator deferiprone (DFP)) iron overload are available, but because retrospective studies to date only identify a correlative, not a causal, relationship between iron overload and reduced survival, there is little consensus on whether benefits of its diagnosis and treatment outweigh risks in this patient population. We aimed to evaluate and characterize the effect of iron loading and iron chelation on ineffective hematopoiesis in NUP98-HOXD13 transgenic (NHD13) mice, a highly penetrant model with peripheral blood cytopenias and ineffective hematopoiesis with dysplasia, consistent with MDS in humans [Lin Blood 2005]. We hypothesize that iron overload in MDS has deleterious biological effects on hematopoiesis with increased likelihood of worsening disease, reversible with iron chelation therapy. Our preliminary data demonstrates that 1) DFP enhances erythroid differentiation in CD34+ cells from MDS patients; and 2) exogenous iron suppresses erythropoiesis in wild type (WT) mice. The current experiments reveal the effect of exogenous iron and DFP in NHD13 mice. Age and gender matched 5 month old mice, 5-10 mice per group, were injected with 20mg iron dextran and/or treated with DFP (175 mg/mouse) in the drinking water over 4 weeks. Treated mice were analyzed and compared with PBS injected NHD13 mice and WT controls. Our experiments demonstrate that NHD13 mice exhibit anemia and leukopenia, increased serum erythropoietin (Epo) concentration, and higher MCV despite no difference in reticulocyte count relative to WT controls (Table I) and no splenomegaly (Fig 1a). In addition, erythroblast ROS concentration (Fig 1b) and iron load in the liver (Fig 1c) increase without differences in transferrin saturation or hepcidin expression in NHD13 relative to WT mice. These findings demonstrate characteristics consistent with human MDS patients. We then analyzed the effects of additional iron loading, iron chelation, or a combination of both on parameters of iron metabolism and erythropoiesis in NHD13 mice. Iron-treated NHD13 mice exhibit higher WBC count, RBC count, and hemoglobin, both DFP- and iron-treated NHD13 mice exhibit decreased serum Epo, and DFP+iron-treated NHD13 mice exhibit decreased serum Epo while increasing RBC count and hemoglobin (Table I). These findings suggest that exogenous iron increases Epo responsiveness and extramedullary erythroid mass in NHD13 mice in a manner similar to what we observed in iron-treated thalassemic mice [Ginzburg Exp Hem 2009]. Furthermore, spleen size increased (Fig 1a) and erythroblast ROS decreased (Fig 1b) only in iron-treated NHD13 mice. These findings suggest that erythroblast ROS is unrelated to excess iron in NHD13 mice. Liver iron increased in iron-treated and decreased in DFP-treated NHD13 mice (Fig 1c) as expected. In addition, CD71 (TfR1) surface expression on bone marrow erythroblasts is suppressed in all treated NHD13 mice but only significantly decreased in DFP-treated mice (Fig 1d), suggesting expected iron restriction. Lastly, DFP- and DFP+iron-treated (but not iron-treated) NHD13 mice decreased the proportion of bone marrow erythroblasts (Fig 1e) and increased erythroid differentiation in NHD13 mouse bone marrow (Fig 1f). Taken together, our findings demonstrate the robustness of NHD13 mice as a model of MDS to study erythropoiesis, the utility of iron injection in NHD13 mice to mimic robust iron overload in MDS, and the effectiveness of DFP in enhancing erythroid differentiation, reversal of erythroid expansion, and Epo responsiveness in NHD13 mice. Additional experiments (i.e. RBC survival, analysis of bone marrow signaling, erythroferrone expression, and parameters of the iron restriction response) to further explore the dysregulation of iron metabolism in NHD13 mice are ongoing.
No relevant conflicts of interest to declare.

This feature is available to Subscribers Only
Sign In or Create an Account Close Modal