

In this issue of Blood, report that increased expression of factor VIII (FVIII) can activate the endoplasmic reticulum (ER) stress response through the formation of amyloid-like fibrils that prevent the further trafficking and secretion of the protein. This study provides insights into the factors that influence the effective secretion of FVIII that may have implications for in vitro protein production systems and in vivo gene therapy.1
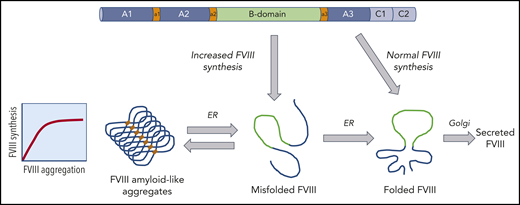
Model for the intracellular trafficking of FVIII that can result in the formation of aggregates. The FVIII polypeptide chain enters the ER where chaperone proteins facilitate folding and stabilize FVIII. In the setting of increased FVIII expression, FVIII can misfold and form aggregates. The formation of these amyloid-like aggregates may be reversible under specific conditions.
Model for the intracellular trafficking of FVIII that can result in the formation of aggregates. The FVIII polypeptide chain enters the ER where chaperone proteins facilitate folding and stabilize FVIII. In the setting of increased FVIII expression, FVIII can misfold and form aggregates. The formation of these amyloid-like aggregates may be reversible under specific conditions.
FVIII deficiency causes the bleeding disorder hemophilia A. It has been known for decades that FVIII is a difficult protein to express both in vitro and in vivo. The first observations were made in vitro in the setting of recombinant protein production. Stable mammalian cell lines (such as Chinese hamster ovary cells, baby hamster kidney cells) were established to synthesize FVIII for the purpose of recombinant protein production used to treat hemophilia A patients. However, the yield of FVIII from these cells was low due to poor expression of the protein. Studies suggested that this low yield may be due to low FVIII messenger RNA levels or poor secretion of FVIII.2
FVIII circulates in the plasma at a concentration of 0.1 to 0.2 μg/mL, which is the lowest concentration of all coagulation factors by at least a 10-fold margin. Early work by Kaufman and colleagues compared FVIII to factor V (FV), which has a similar domain structure and protein size. Even in the setting of identical in vitro expression systems, FV was expressed at 5- to 10-fold higher levels than FVIII, suggesting a unique feature of FVIII that prevents effective expression of the protein.3
Studies to investigate the cellular processing of FVIII revealed that high-level expression of FVIII can cause ER stress and activate the unfolded protein response (UPR).4 The UPR is a strategy used by the cells to adapt to cellular stress; however, under conditions of prolonged activation, the cell may undergo apoptosis. In the current study by Poothong et al, several factors that influence FVIII protein misfolding and activation of the UPR pathways are uncovered. When FVIII synthesis is increased or glucose metabolism is inhibited, FVIII aggregates into amyloid-like fibrils that accumulate in the ER. The study also demonstrates that a specific region of the FVIII A1 domain appears to be responsible for the initiation of the amyloid formation. Further analysis comparing FV with FVIII pinpointed specific residues that impact the aggregation and secretion of FVIII. Importantly, the chaperone protein BiP bound to this motif in the A1 domain and prevented aggregation of FVIII. However, under conditions of cellular stress such as increased FVIII expression, FVIII may accumulate in the ER, which may saturate a BiP-dependent mechanism that prevents aggregation. Interestingly, it appears that this may be a dynamic process that could be reversible (see figure).
Although the focus of the current studies is in vitro, the findings may have significant implications for clinical studies of gene therapy for hemophilia A. In both hemophilia A mouse and dog preclinical models, it is challenging to express therapeutic levels of FVIII after adeno-associated viral vector (AAV)-mediated gene delivery compared with AAV delivery of factor IX. Current approaches to express FVIII use robust hepatocyte-specific promoter elements and codon-optimized gene sequences in an effort to increase FVIII expression. In comparison with the first-generation FVIII transgene constructs, this has led to an increase in FVIII expression per transgene, which increases the FVIII load per cell. In addition, because the AAV vector transduction may result in different numbers of vectors delivered per cell, the levels of FVIII expression per cell is likely heterogeneous. However, careful evaluation of the FVIII distribution throughout the liver tissue after AAV gene delivery has not been performed. The ongoing clinical studies for AAV delivery of FVIII show promising efficacy data with a median FVIII level of 60% of normal activity (range 11% to 95%) at 1 year after vector administration with no major safety concerns; however, there was a loss of FVIII expression in the majority of the subjects (10 of 13 subjects) that occurred after the first year.5 This was not observed in preclinical studies in hemophilia A dogs that were followed for up to a decade; however, these dogs were treated with first-generation FVIII transgene constructs that did not express FVIII at high levels and resulted in more modest therapeutic FVIII levels of <10% of normal levels.6,7 It is unclear if the total FVIII expression level or the expression on a cellular basis is an important consideration for gene-based therapeutics. The studies presented here suggest that 1 hypothesis for the decline in FVIII expression could be that high-level FVIII expression within a cell results in the accumulation of FVIII aggregates that leads to a cellular stress response. It is also important to note that current clinical gene therapy efforts target expression to hepatocytes that are not the native site of FVIII expression, and it remains unknown if there may be differences in the cellular stress response to FVIII in the endogenous site of FVIII synthesis, the liver sinusoidal endothelial cell.8,9
One of the outstanding questions is to understand if this phenomenon indeed occurs in vivo. Although it is possible that the secretion of FVIII is impaired due to aggregation in vivo, it will be important to understand if this affects the outcome in the setting of gene therapy. Although the in vitro data provided by Poothong et al suggest that FVIII aggregation is dependent on the FVIII expression levels, it will be essential, yet more challenging, to understand if this dose-dependent effect occurs in vivo. Moreover, it will be important to determine if this bottleneck can be overcome in the setting of gene delivery. Modifications of key residues in the FVIII protein may prevent aggregation and improve FVIII secretion. Several mutations of FVIII based on the FV amino acid sequence (F309S and L303E/F309S) showed reduced aggregation and increased FVIII secretion in this study. These and other variants of FVIII that result in improved expression may provide a strategy to improve the cellular trafficking of FVIII and avoid activation of the UPR, which may impact the outcomes of gene therapy.
Conflict-of-interest disclosure: D.E.S. received royalties from Spark Therapeutics.
