

Platelet activation supports procoagulant activity through phosphatidylserine exposure, secretion of procoagulant factors, and receptor conformational change. For example, thrombin-stimulated platelets bind factor VIII (fVIII) via a macromolecular complex including oligomeric fibrin and the active αIIbβ3 receptor (Phillips et al, JTH 2004; Gilbert et al, Blood 2015). Thus, coagulation assays in which phospholipid vesicles are substituted for platelets do not fully emulate modulators of fVIII activity. Indeed, inhibition of platelet-supported fVIII activity by a panel of mAbs against the C2 domain was not correlated to inhibition of vesicle-dependent activity (Chatterjee et al, JTH 2020). An obstacle to adoption of a platelet-based assay for fVIII is the need for fresh platelets. Therefore we asked whether cryopreserved platelets might support fVIII activity similarly to fresh platelets.
Apheresis platelets were mixed with cryopreservatives with or without calcium chelators, in various aliquot sizes, and frozen on various cooling media. Cryopreserved platelets were compared to non-preserved apheresis platelets with regard to agonist response and support of procoagulant activity. Cryopreservation resulted in an increase in subcellular debris and an unresponsive fraction of platelets with decreased forward scatter judged by flow cytometry. Optimized results were obtained when platelet rich plasma with 5% DMSO, in 1 mL aliquots was frozen on powdered dry ice, and stored at -150C. Purification of thawed platelets using a density gradient removed debris and decreased unresponsive platelets resulting in a forward and side scatter profile comparable to fresh platelets. We refer to these as cryopreserved platelets (CryoPlts).
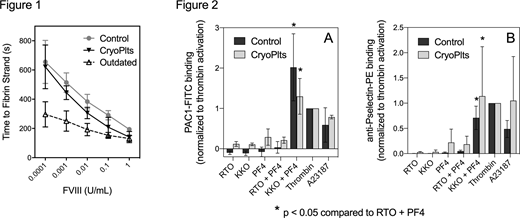
CryoPlts were compared to control and outdated apheresis platelets. As with fresh platelets, procoagulant activity of CryoPlts increased with thrombin receptor agonist peptides (TRAP) 1 & 4 and supported a log-linear relationship between time to initial fibrin strand formation and fVIII activity over a range of 0.0001 - 1 u/mL (Fig 1). Further, the degree of inhibition of fVIII activity by mAbs ESH4 and G99 against the fVIII C2 domain, was the same on control and CryoPlts, but markedly different from inhibition in an aPTT-based inhibitor assay. In contrast, outdated apheresis platelets had increased procoagulant activity, minimal agonist response and a shallow curve with varying fVIII concentration. Flow cytometry studies with lactadherin-FITC indicated that 33 ± 14% of CryoPlts had high PS exposure, and the size of this population was minimally affected by TRAP 1+4. In contrast, the main platelet population had a small, uniform, increment in PS exposure comparable to control platelets. Surprisingly, the PS-rich platelets did not significantly affect the time to fibrin formation, confirming that the viable platelets, with limited PS exposure, provide much of the support for fVIII-related procoagulant activity. Flow cytometry indicated αIIbβ3 activation (PAC1-FITC) and α-granule release (anti-P-selectin-PE) were qualitatively intact on CryoPlts, although staining was decreased 70% for PAC1 and 57% for Psel.
We also tested whether CryoPlts may be utilized for evaluating response to anti-PF4-heparin antibodies, relevant to heparin-induced thrombocytopenia (HIT). We evaluated platelet response to platelet factor 4 (PF4) and a platelet-activating anti-PF4 antibody (KKO), a combination that induces activation similar to authentic autoimmune antibodies for HIT. The non-activating PF4 antibody RTO served as a negative control. Geometric mean response, corrected for background, was normalized to response to thrombin activation. Both fresh and CryoPlts responded with increases in PAC1 (Fig 2A) and anti-Psel (Fig 2B) binding in response to KKO/PF4 compared to RTO/PF4 . This data demonstrates that the qualitative αIIbβ3 and P-selectin response to HIT-like antibodies is intact.
Our results demonstrate a refined cryopreservation protocol of apheresis platelets. These platelets maintain qualitative agonist responsiveness with near-normal support for factor VIII activity, suggesting that they could be used for other platelet-based laboratory or diagnostic assays. Further, our results suggest that major procoagulant activity is provided by platelets with very limited PS exposure, an area for further investigation.
No relevant conflicts of interest to declare.

This feature is available to Subscribers Only
Sign In or Create an Account Close Modal