

Key Points
Constitutive activation of NF-κB pathways early in B-cell development alters the transcriptional program and loses B cells via apoptosis.

Abstract
There is a considerable body of work exploring the role of NF-κB family of transcription factors in the maturation and functions of later stage B cells; however, their role in the earlier bone marrow stages of development is less well understood despite the demonstration that NF-κB activity is present at all early stages of B-cell development. To explore the consequences of early, B cell–targeted constitutive activation of both NF-κB pathways on B-cell development, we generated mice that have either or both. NF-κB pathways constitutively activated beginning in early pro-B cells. In marked contrast to activating a single pathway, we found mice with both pathways constitutively activated displayed a profound loss of B cells, starting with early pro-B cells and peaking at the late pro-B-cell stage, at least in part as a result of increased apoptosis. This effect was found to be cell autonomous and to have striking phenotypic consequences on the secondary lymphoid organs and circulating antibody levels. This effect was also found to be temporal in nature as similar activation under a Cre expressed later in development did not result in generation of a similar phenotype. Taken together, these findings help to shed further light on the need for tight regulation of the NF-κB family of transcription factors during the various stages of B-cell development in the bone marrow.
Introduction
The development of B cells from hematopoietic stem cells in the bone marrow is a sequential process that can be classified into stages based on the expression of cell surface molecules.1 The earliest committed B-cell progenitors are pre-pro B cells, the first population to express the B cell–restricted surface marker B220/CD45R. Subsequently, cells express first a pre-B-cell receptor (BCR), then undergo VD(J) recombination to generate the unique heavy and light chains of the BCR. If this rearrangement is successful, the new B cells egress into the periphery to finish maturing.1
The NF-κB family of transcription factors are involved in the maturation, survival, and antigen response of B cells, including the response to the survival factor BAFF and BCR signaling.2 The NF-κB family consists of 5 members: RelA (p65), RelB, and c-Rel, which are synthesized in mature form and interact in the cytoplasm with inhibitors until activated by an appropriate signal, and NF-κB1 (p105/p50) and NF-κB2 (p100/p52), which are synthesized as precursor molecules and catalytically processed to an active form. NF-κB family members then translocate to the nucleus as various homo- and heterodimers to induce gene transcription.2 Although a considerable body of work explores the role of NF-κB in the maturation and functions of peripheral B cells, the role of NF-κB in earlier bone marrow development is less well understood, despite the demonstration that NF-κB activity is present at all early development stages, with a peak in activity in cells expressing the pre-BCR.3
The role NF-κB plays in early B-cell development has been investigated through many single-gene knockout mice, but B-cell development in bone marrow was not measurably affected, possibly because of redundancies in the signaling pathways or NF-κB subunits.4-12 Attempts have been made to address this through the use of more sophisticated genetic approaches that target both pathways or 2 or more subunits simultaneously, including mb1-cre targeted deletions of NEMO, IKK1, IKK2, and others.10,13-15
All found development to be impaired at the pre-B and immature B-cell stages and reduced numbers of mature B cells in the periphery. Pro-B cells were reduced in mice expressing a dominant negative form of IκBα.16 Impairment of both NF-κB pathways using a knock-in of a kinase dead form of IKKα resulted in reductions in the earliest stage B cells among other defects, likely because of the altered expression of Pax5 and IRF4.16 Paradoxically, mice lacking the p100 inhibitor, causing constitutive activation of the alternative NF-κB pathway, also had reduced expression of Pax5, which was found to be responsible for the arrested transition from pre-pro-B to pro-B cells.17 In contrast, constitutive activation of the alternative NF-κB pathway in CD19Traf3−/− mice18 and of the classical NF-κB pathway in CD19IKK2ca mice12 was found to have no impact on the development of B cells in the bone marrow, despite the increased accumulation of mature B cells in the periphery.
The majority of studies to date have focused on the inactivation of the classical or alternative pathways alone or together. We wanted to explore the consequences of early, B cell–targeted constitutive activation of both NF-κB pathways on B-cell development, in part because several peripheral B-cell tumors, such as mucosa-associated lymphoid tissue lymphoma19 and multiple myeloma,20 are known to depend on such activation. We generated such mice with either or both NF-κB pathways constitutively activated beginning in early pro-B cells. In marked contrast to studies activating a single pathway and contrary to the expectation of greatly enhanced numbers of B cells, we found these mice displayed a profound loss of B cells, starting with early pro-B cells and peaking at the late pro-B-cell stage as a result of increased apoptosis. This effect was found to be cell autonomous and to have striking phenotypic consequences on the secondary lymphoid organs. B cells from these mice had altered transcriptional profiles as well as changes in their ability to respond to stimuli. This effect was also found to be temporal in nature as the same constitutive activation under control of CD21 cre21 resulted in no impairment. Taken together, these findings help to shed further light on the need for tight temporal regulation of the NF-κB family of transcription factors in the development of B cells in the bone marrow.
Methods
Mice
Traf3flx/flx conditional knockout mice were generated as described (supplemental Figure 1). IKK2CA transgenic, CD45.1, mb1 cre, and CD21 cre mice were purchased from Jackson Laboratories (Ellsworth, ME). Mice were analyzed at 8 to 12 weeks of age. Mice were bred and housed in National Institute of Allergy and Infectious Diseases facilities, and all experiments were done with the approval of the National Institute of Allergy and Infectious Diseases Animal Care and Use Committee and in accordance with all relevant institutional guidelines.
Antibodies and flow cytometry
The following antibodies were used for flow cytometric analysis: B220 (RA3-6B2), CD19 (1D3), Ly51/BP-1 (6C3), TCRb (H57-597) and CD3 (145-2C11) (eBioscience, San Diego, CA); CD43 (1B11) and IgD (11-26c.2a) (Biolegend, San Diego, CA), and IgM (R6-60.2), CD24, (M1/69) and CD45.2 (104) (BD Biosciences San Jose, CA). Flow cytometry was performed using a Canto II (BD Biosciences) and data were analyzed using FlowJo software. Caspase 3/7 was detected using the Magic Red Caspase 3 and 7 assay kit (Immunochemistry Technologies, Bloomington, MN).
Cell purification
B220+ cells were purified from bone marrow after the generation of single-cell suspensions and red blood cell lysis using ACK lysis buffer (Lonza, Gaithersburg, MD). Cells were incubated with B220 beads (Miltenyi Biotec, San Diego, CA) and B220+ cells selected using an AutoMACS (Miltenyi Biotec).
Purity was measured by flow cytometry. For flow sorting, the enriched bone marrow B220+ cells were then labeled with B220, CD43, Ly51, and CD24 and then sorted for groups A, B, and C using a FACS Aria (Becton Dickinson, San Jose, CA).
Real-time polymerase chain reaction
RNA was extracted from sorted cells using the RNEasy Mini Kit (Qiagen), complementary DNA synthesized using the Qantitect RT kit (Qiagen, Germantown, MD), and gene expression analyzed TaqMan Gene Expression Assay kits (Thermo Fisher Scientific, Waltham, MA).
Bone marrow culture
Purified B220+ cells were cultured in RPMI supplemented with 10% fetal bovine serum, nonessential amino acids, sodium pyruvate, and β mercaptoethanol (complete RPMI) at a concentration of 3 × 106 cells/well in the presence of 20 ng/mL interleukin-7 (IL-7; R&D Systems, Minneapolis, MN) for 4 days. For further B-cell development, cells were washed and cultured for an additional 3 days in the complete RPMI. Cells were analyzed by flow cytometry on days 4 and 7 of culture.
Proliferation assay
Purified B220+ cells were labeled with CellTrace Violet (Invitrogen, Carlsbad, CA) and then seeded at 1.5 × 106 cells/well in complete RPMI supplemented with 20 ng/mL IL-4 (R&D Systems). Cells were cultured for 4 days with media alone or in the presence of 100 ng/mL lipopolysaccharides (LPS; Sigma Aldrich, Burlington, MA), 10 ∝g/mL anti-IgM (BD), 2 ∝g/mL anti-CD40 (BD), or 10 ng/mL phorbol 12-myristate 13-acetate plus 500 ng/mL Ionomycin (Sigma Aldrich). Cells were then washed, stained with live/dead Aqua (Invitrogen), and analyzed by flow cytometry.
Histology
Spleens were fixed in 4% paraformaldehyde and paraffin embedded. Sections stained with hematoxylin and eosin and visualized on an Olympus BX50 microscope.
Bone marrow transplantation
CD45.1 mice received 900 rads of γ irradiation and 24 hours later given donor bone marrow cells. Briefly, donor bone marrow was isolated from the femurs and tibias of CD45.1 wild-type (WT), CD45.2 WT, or CD45.2 double-pathway (DPWY) mice. Mice received a mixture of 1 × 106 CD45.1 cells with either 1 × 106 CD45.2 WT or DPWY cells (a total of 2 × 106 cells) by intravenous injection. The bone marrow of recipient mice was analyzed by flow cytometry 8 weeks after transfer.
Serum immunoglobulin and anti-dsDNA ELISA
Serum immunoglobin levels were measured using Immulon 4HBX plates (Thermo Scientific) coated with 5 μg/mL of anti-immunoglobulin G (IgG; Southern Biotech). After incubation with mouse sera, antibodies to IgM, IgG, IgA, or IgG2c conjugated with biotin (Southern Biotech) were added, followed with streptavidin-horseradish peroxidase/trinitrophenyl detection. Mouse double-stranded DNA (dsDNA) IgG was detected by enzyme-linked immunosorbent assay (ELISA) using the dsDNA IgG kit (5120) (α Diagnostic International, San Antonio, TX).
Results
Early constitutive activation of both NF-κB pathways results in a systemic loss of B cells
We examined the impact of a targeted early constitutive activation of both NF-κB pathways in developing B cells, something that has not previously been addressed. We generated Traf3flx/flx mice to allow for cell-specific deletion of Traf3, leading to constitutive activation of the alternative NF-κB (supplemental Figure 1).
These were crossed with IKK2ca animals12 and we then generated mice in which either the classical, alternative, or both NF-κB pathways (hereafter referred to as DPWY) were constitutively activated under the control of the B cell–specific promoter, mb1, which is first expressed in early pro-B cells. Constitutive activation of either classical (class), and particularly the alternative (alt) pathways singly caused significant hyperplasia of B cells in spleens (Figure 1A-C), including an accumulation of marginal zone B cells (data not shown), a finding consistent with published data for CD19 cre.12,18 These changes were also seen at the gross histological level, with spleens displaying enlarged B-cell follicles and wider marginal zones (supplemental Figure 2). It was therefore surprising that mice with both pathways constitutively activated (mb1DPWY) did not develop B-cell hyperplasia, but instead displayed a dramatic decrease in B cells in the spleen (Figure 1; littermate controls [LMC]) and all other lymphoid tissues examined (supplemental Figure 2A). These data were surprising given the contrast to single pathway activation and the systemic nature of the effect led us to examine the bone marrow of these mice.
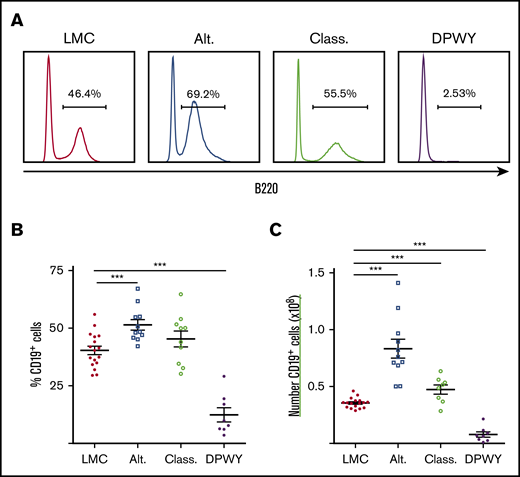
Changes in B cells following activation of NF-κB pathways. (A) Representative histograms of the percentage of B220+ cells in the spleens of mice with the classical (Class.), alternative (Alt.), or both NF-κB pathways (DPWY) constitutively activated under the control of mb1 cre compared with LMC. (B) Summary data of the proportion of B cells in the spleen. (C) Summary data of the number of B cells in the spleen. Data for A-C are from n = 8-17 mice per group, from 3-5 independent experiments; summary data are shown as the mean ± standard error of the mean (SEM). ***P < .001.
Changes in B cells following activation of NF-κB pathways. (A) Representative histograms of the percentage of B220+ cells in the spleens of mice with the classical (Class.), alternative (Alt.), or both NF-κB pathways (DPWY) constitutively activated under the control of mb1 cre compared with LMC. (B) Summary data of the proportion of B cells in the spleen. (C) Summary data of the number of B cells in the spleen. Data for A-C are from n = 8-17 mice per group, from 3-5 independent experiments; summary data are shown as the mean ± standard error of the mean (SEM). ***P < .001.
Loss of B cells occurs early in development
Because the loss of B cells in mb1DPWY mice included the bone marrow (supplemental Figure 3A), we wanted to more closely investigate whether there was a particular stage of B-cell development being affected. Expression of the mb1 gene (CD79a/Igα) begins in early pro-B cells; by the late pro-B-cell stage, the majority of cells express this protein as part of the pre-BCR22 (supplemental Figure 4). Using staining based on the Hardy classification system,1 we carefully examined the different B-cell developmental stages and found that not only were there proportionally fewer overall B cells in the bone marrow of mb1DPWY mice (Figure 2A top, quantitated in Figure 2B), very few cells were present from the late pro-B-cell stage/fraction C; indicating a developmental arrest at the early pro-B cells/fraction B stage (Figure 2A,D). The effects of the dual pathway activation are even more striking when the overall impact within the bone marrow is taken into account, not only are the fewer total B cells (Figure 2B), an absence of immature and recirculating cells is reflected in the significant reduction of B220+ cells that downregulate CD43 and a reciprocal accumulation of CD43-expressing cells (Figure 2C). To quantify the impact on the different developmental stages, the relative proportion of each subset was calculated as a proportion of bone marrow cells, allowing us to account for a significantly reduced B-cell population, changes in CD43 expression within the B220+ population, and changes in the proportions of each developmental stage. This analysis showed that, other than fractions A and B, all subsets were significantly reduced (Figure 2D), indicating that significantly fewer cells were progressing through the development pathway, although the few cells that appeared to progress beyond the pro-B-cell stage did migrate appropriately to the various secondary lymphoid organs (Figure 2; supplemental Figure A).
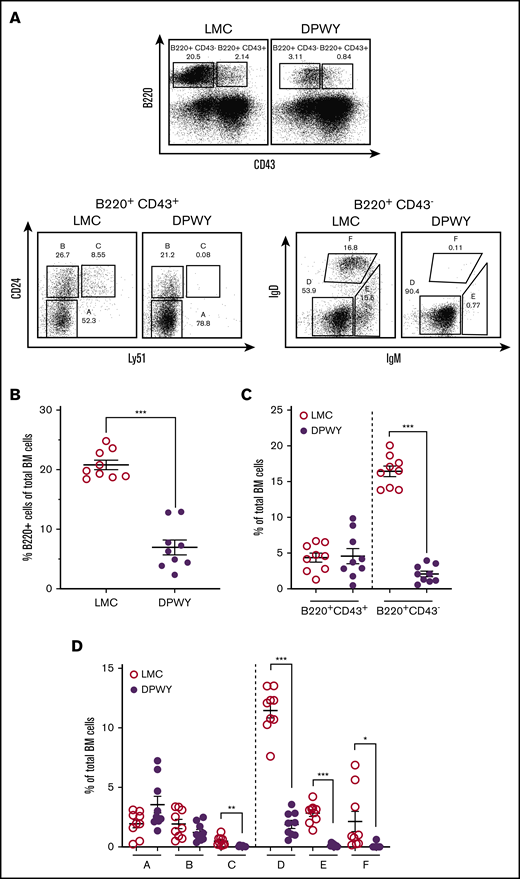
B-cell changes begin early in bone marrow development. (A) Flow cytometry staining of bone marrow B cells is based on the Hardy classification system and was conducted as described in the “Methods” section, cells are gated on live lymphocytes for analysis. Representative dot plots and summary data are shown for LMC and mb1DPWY bone marrow. (B) Summary data for the percentage of B220+ cells in total bone marrow. (C) The percentage of CD43− vs CD43+ B220 expressing cells in the total bone marrow of mb1DPWY vs LMC showing proportionally fewer more mature (CD43−) B cells present in the bone marrow of mb1DPWY mice. (D) The percentage of each developmental subset (A-F) in the total bone marrow of mb1DPWY vs LMC showing the significant loss of B cells in fraction C onward. Cells are gated on live lymphocytes for analysis. Summary data of bone marrow subsets are for n = 9 mb1DPWY and n = 9 LMC from 3 independent experiments; data are shown as the mean ± SEM. *P < .05, **P < .01, ***P < .001.
B-cell changes begin early in bone marrow development. (A) Flow cytometry staining of bone marrow B cells is based on the Hardy classification system and was conducted as described in the “Methods” section, cells are gated on live lymphocytes for analysis. Representative dot plots and summary data are shown for LMC and mb1DPWY bone marrow. (B) Summary data for the percentage of B220+ cells in total bone marrow. (C) The percentage of CD43− vs CD43+ B220 expressing cells in the total bone marrow of mb1DPWY vs LMC showing proportionally fewer more mature (CD43−) B cells present in the bone marrow of mb1DPWY mice. (D) The percentage of each developmental subset (A-F) in the total bone marrow of mb1DPWY vs LMC showing the significant loss of B cells in fraction C onward. Cells are gated on live lymphocytes for analysis. Summary data of bone marrow subsets are for n = 9 mb1DPWY and n = 9 LMC from 3 independent experiments; data are shown as the mean ± SEM. *P < .05, **P < .01, ***P < .001.
The role of the NF-κB pathways is B cell–autonomous
To determine whether deletion of B cells at the late pro-B-cell stage is directly the result of cell-autonomous dysregulation of NF-κB signaling, rather than a lack of factors produced by the B cells themselves acting directly or indirectly on B cells, we performed a bone marrow cotransfer; irradiated congenic WT mice received a transfer of either mixed congenic WT CD45.1 and LMC CD45.2 or a combination WT (CD45.1) and mb1DPWY (CD45.2) bone marrow cells. Although CD45.2 LMC B cells developed normally (Figure 3A-B), we found that the B-cell development phenotype seen in mb1DPWY mice was recapitulated in the transferred bone marrow cells. We observed fewer B220+ cells (Figure 3B), fewer CD43− cells (Figure 3C), and significantly fewer cells developing beyond fraction B/late pro-B cells, as described in the previous section (Figure 3A,D). This occurred independently of the presence of WT stroma and the presence of normally developing WT B cells, ruling out extrinsic deficiencies. Furthermore, the same phenotype occurred in an in vitro culture system where the requirement for bone marrow stromal cells is removed. We found that very few cells progressed beyond the late pro-B-cell stage in culture at day 4 (Figure 3E-F). Taken together, these results clearly suggest a cell autonomous role for NF-κB signaling at the early pro-B-cell stage of development.
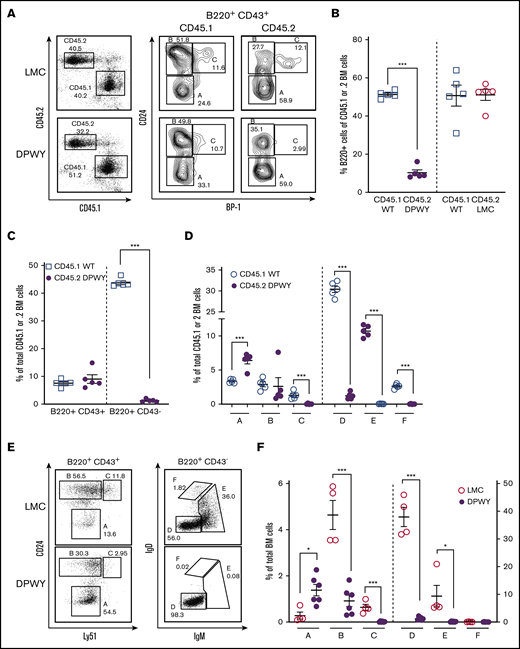
The bone marrow developmental issue is B cell–intrinsic. (A) Flow cytometric analysis of lethally irradiated CD45.1 recipient mice receiving equal numbers of bone marrow cells (1 × 106 cells/genotype) from CD45.2 LMC and CD45.2 mb1DPWY by IV injection. Bone marrow was analyzed by flow cytometry based on the Hardy classification system 8 weeks after transfer. Data are shown as a representative plot for n = 5 mice per group from a single experiment. Summary data for the percentage of B220+ in (B) total bone marrow, the percentage of B220+ cells maturing B cells as measured by CD43 expression is decreased in CD45.2 mb1DPWY compared with (C) CD45.1 WT cotransferred cells, and the percentage of each (A-F) developmental subset in the total bone marrow of CD45.2 mb1DPWY vs (D) CD45.1 WT cells showing the significant loss of B cells in fraction C onward. Data are shown as mean ± SEM. (E) Representative plots and (F) summary data of B-cell developmental stages in day 4 in vitro bone marrow. Equal numbers of total bone marrow cells from mb1DPWY and LMC mice were cultured for 4 days in the presence of IL-7 and harvested for analysis. Cells were analyzed by flow cytometry based on the Hardy classification system. Data are representative of n = 1-3 mice/genotype from 2 independent experiments. *P < .05, ***P < .001.
The bone marrow developmental issue is B cell–intrinsic. (A) Flow cytometric analysis of lethally irradiated CD45.1 recipient mice receiving equal numbers of bone marrow cells (1 × 106 cells/genotype) from CD45.2 LMC and CD45.2 mb1DPWY by IV injection. Bone marrow was analyzed by flow cytometry based on the Hardy classification system 8 weeks after transfer. Data are shown as a representative plot for n = 5 mice per group from a single experiment. Summary data for the percentage of B220+ in (B) total bone marrow, the percentage of B220+ cells maturing B cells as measured by CD43 expression is decreased in CD45.2 mb1DPWY compared with (C) CD45.1 WT cotransferred cells, and the percentage of each (A-F) developmental subset in the total bone marrow of CD45.2 mb1DPWY vs (D) CD45.1 WT cells showing the significant loss of B cells in fraction C onward. Data are shown as mean ± SEM. (E) Representative plots and (F) summary data of B-cell developmental stages in day 4 in vitro bone marrow. Equal numbers of total bone marrow cells from mb1DPWY and LMC mice were cultured for 4 days in the presence of IL-7 and harvested for analysis. Cells were analyzed by flow cytometry based on the Hardy classification system. Data are representative of n = 1-3 mice/genotype from 2 independent experiments. *P < .05, ***P < .001.
Loss of B cells is due to apoptosis and surviving cells are of immature phenotype
To determine whether this striking loss of cells was due to apoptosis, we measured caspase 3 and 7 activity in both mb1DPWY and LMC bone marrow cells and found that there is indeed a significant increase in caspase activity in late pro-B/fraction C cells of mb1DPWY mice (Figure 4A-B). This trend begins in early pro-B/fraction B cells and corresponds closely with the expression pattern of mb1 (supplemental Figure 4), suggesting a direct role for the NF-κB pathways at this stage in B-cell development. Analysis of messenger RNA levels of different genes related to apoptosis in these cells showed higher expression of pro-apoptotic genes such as Bim and Bax and reduced expression of anti-apoptotic genes such as Bcl-2 and Bcl-XL (supplemental Figure 5). Interestingly, a second peak in caspase 3 and 7 activity was observed in immature B cells/fraction E, and borne out with cell proportions in Figure 2 and3, suggesting those few cells passing through the late pro-B/fraction C stage subsequently have defects causing deletion before egress from the bone marrow, resulting in even fewer B cells in the periphery. Those very rare cells able to escape deletion and egress to the periphery were found to exhibit a less mature phenotype as measured by expression of IgD when compared with LMC (Figure 4C) and the ratio of mature vs immature cells was found to be significantly lower, suggesting that these cells would be less likely to be fully functional in an immune response (Figure 4D). In keeping with these cellular findings, quantitative polymerase chain reaction analysis of bone marrow fraction B/early pro-B cells showed significantly reduced expression of transcription factors critical for B-cell lineage commitment and development, Ebf1, Pax5, and Ikaros, indicating that the altered activation of NF-κB pathways has profound effects on the transcriptional program of developing B cells (Figure 4E).

B cells are deleted early in development because of NF-κB activation and escaping cells are immature. (A) Representative plots for Caspase 3 and 7 expression in development stage C and (B) summary data for Caspase 3 and 7 expression in different bone marrow B-cell development stages. Cells are gated on live B220+ cells and then subclassified using the Hardy system before Caspase 3 and 7 expression was assessed as a percentage in each developmental stage. Data are from n = 9 mice/genotype, shown as the mean ± SEM and were collected in 3 independent experiments. (C) Representative plots for mature and immature B cells in the spleen of LMC and mb1DPWY mice. Cells are gated on live B220+ cells and stained for surface IgM and IgD, with immature cells being IgDlo/− and mature cells being IgD+. (D) Summary data of the ratio of mature to immature B cells in the spleen of WT and mb1DPWY mice, data are from n = 11 WT and n = 7 mb1DPWY mice, shown as the mean ± SEM and were collected in 4 independent experiments. (E) Gene expression of Pax5, Ebf1, and Ikaros in Hardy group B cells, data are from n = 3 replicates/genotype (pooled from 2-3 mice/replicate). Data shown as fold induction compared with LMC controls. *P < .05, **P < .01, ***P < .001.
B cells are deleted early in development because of NF-κB activation and escaping cells are immature. (A) Representative plots for Caspase 3 and 7 expression in development stage C and (B) summary data for Caspase 3 and 7 expression in different bone marrow B-cell development stages. Cells are gated on live B220+ cells and then subclassified using the Hardy system before Caspase 3 and 7 expression was assessed as a percentage in each developmental stage. Data are from n = 9 mice/genotype, shown as the mean ± SEM and were collected in 3 independent experiments. (C) Representative plots for mature and immature B cells in the spleen of LMC and mb1DPWY mice. Cells are gated on live B220+ cells and stained for surface IgM and IgD, with immature cells being IgDlo/− and mature cells being IgD+. (D) Summary data of the ratio of mature to immature B cells in the spleen of WT and mb1DPWY mice, data are from n = 11 WT and n = 7 mb1DPWY mice, shown as the mean ± SEM and were collected in 4 independent experiments. (E) Gene expression of Pax5, Ebf1, and Ikaros in Hardy group B cells, data are from n = 3 replicates/genotype (pooled from 2-3 mice/replicate). Data shown as fold induction compared with LMC controls. *P < .05, **P < .01, ***P < .001.
Temporal activation of the NF-κB pathways is critical for phenotype
A major issue we wished to address was whether the timing of NF-κB activation was leading to the observed loss of B cells. To address this, we generated CD21DPWY mice; CD21 is expressed from the transitional B-cell stage onwards, after B cells have egressed from the bone marrow and therefore considerably later in the cell development pathway. This allowed for a larger temporal separation than CD19 Cre would have provided. The CD21DPWY mice did not exhibit B cell loss and instead had a mild hyperplasia at 8 to 12 weeks of age (Figure 5A-B), resulting in larger spleens and larger B-cell follicles when compared with both control animals and the reduced B-cell follicles observed in mb1DPWY spleens (data not shown). B cells were not deleted in all lymphoid organs tested (supplemental Figure 2B) and normal development of B cells was observed in the bone marrow of CD21Traf3−/− animals (data not shown), confirming that the effect seen in mb1DPWY is both unique and temporal in nature.
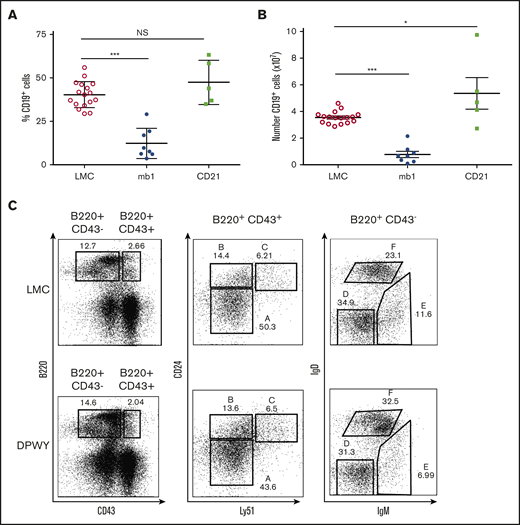
Temporal activation of B cells determines phenotype. Summary data of (A) the proportion of B cells in the spleen and (B) the number of B cells in the spleen of LMC, mb1DPWY, and CD21DPWYmice. Data are from n = 5-17 mice per group and summary data are shown as the mean ± SEM of data collected from 3 independent experiments. (C) Representative plots of bone marrow B-cell development based on the Hardy classification system in LMC and CD21DPWY mice. Data are representative of n = 9 LMC and n = 5 CD21DPWY mice from 3 independent experiments. ***P < .001. NS, not significant.
Temporal activation of B cells determines phenotype. Summary data of (A) the proportion of B cells in the spleen and (B) the number of B cells in the spleen of LMC, mb1DPWY, and CD21DPWYmice. Data are from n = 5-17 mice per group and summary data are shown as the mean ± SEM of data collected from 3 independent experiments. (C) Representative plots of bone marrow B-cell development based on the Hardy classification system in LMC and CD21DPWY mice. Data are representative of n = 9 LMC and n = 5 CD21DPWY mice from 3 independent experiments. ***P < .001. NS, not significant.
Functional defects of mb1DPWY B cells
Given the less mature state of the small number of escaping B cells in the periphery of mb1DPWY mice, we assessed potential functional alterations using a proliferation assay. Purified B220+ bone marrow cells were exposed to different stimuli for 3 days and were found to have significantly diminished proliferative responses to both LPS and anti-CD40 (Figure 6A-C); however, the response to anti-IgM and phorbol 12-myristate 13-acetate/Ionomycin were only slightly reduced compared with controls.
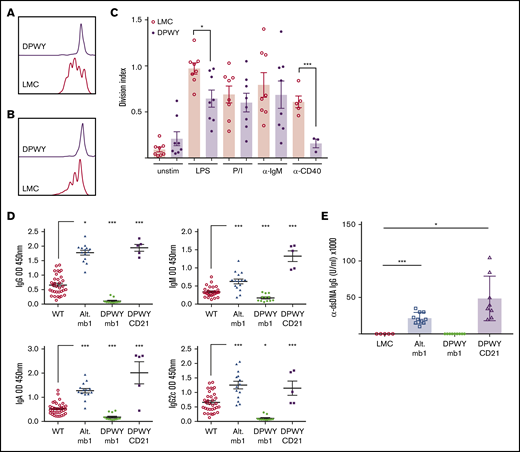
Functional defects of DPWY mb1 B cells. Representative proliferation plots for live B220+ bone marrow B cells stimulated for 72 hours with (A) LPS, (B) anti-CD40 monoclonal antibody, and (C) summary data of the division index of bone marrow B220+ B cells stimulated for 72 hours with the indicated stimuli. Data shown are representative of n = 8 LMC and n = 8 mb1DPWY mice from 4 independent repeats for LPS, anti-IgM, and PI and n = 5 LMC and n = 3 mb1DPWY from 2 independent repeats for anti-CD40. Data are shown as the mean ± SEM. (D) Endogenous levels of serum IgG, IgM, IgA, and IgG2c were measured by ELISA as described in the “Methods.” Data shown are for n = 28 LMC mice, n = 14 mb1Traf3−/−, n = 12 mb1DPWY, and n = 5 CD21DPWY mice. Data are presented as mean ± SEM. (E) Serum levels of anti-dsDNA antibodies were measured at 3 months of age, data are shown as the mean ± SEM for n = 5 LMC, n = 10 alternative pathway, n = 9 mb1DPWY, and n = 8 CD21DPWY mice. *P < .05, ***P < .001.
Functional defects of DPWY mb1 B cells. Representative proliferation plots for live B220+ bone marrow B cells stimulated for 72 hours with (A) LPS, (B) anti-CD40 monoclonal antibody, and (C) summary data of the division index of bone marrow B220+ B cells stimulated for 72 hours with the indicated stimuli. Data shown are representative of n = 8 LMC and n = 8 mb1DPWY mice from 4 independent repeats for LPS, anti-IgM, and PI and n = 5 LMC and n = 3 mb1DPWY from 2 independent repeats for anti-CD40. Data are shown as the mean ± SEM. (D) Endogenous levels of serum IgG, IgM, IgA, and IgG2c were measured by ELISA as described in the “Methods.” Data shown are for n = 28 LMC mice, n = 14 mb1Traf3−/−, n = 12 mb1DPWY, and n = 5 CD21DPWY mice. Data are presented as mean ± SEM. (E) Serum levels of anti-dsDNA antibodies were measured at 3 months of age, data are shown as the mean ± SEM for n = 5 LMC, n = 10 alternative pathway, n = 9 mb1DPWY, and n = 8 CD21DPWY mice. *P < .05, ***P < .001.
It was reported that CD19Traf3−/− mice, which have constitutive activation of the alternative pathway under the control of CD19 cre, develop autoimmune disease, including hypergammaglobulinemia and autoantibody production.18 We therefore examined whether the significant reduction in B-cell number and responses that are observed in mb1DPWY mice impacted immunoglobulin production. To this end, we collected serum from mb1Traf3−/− mice, mb1DPWY, CD21DPWY, and LMC animals and looked at the immunoglobulins present and found that mb1DPWY mice had significantly lower levels of all circulating immunoglobulins as measured by ELISA (Figure 6D). In contrast, mb1Traf3−/− mice developed hypergammaglobulinemia, as was previously reported in CD19Traf3−/− animals, including significantly elevated levels IgG2c, which has been associated with autoimmune disease. Indeed, we found that only mb1Traf3−/− and CD21DPWY mice had detectable levels of anti-dsDNA antibodies present (Figure 6E), displaying signs of autoimmunity that are consistent with those reported by Xie et al.18 These data indicate that the aberrant function of B cells present in mb1DPWY mice do not generate an autoimmune response.
Discussion
B-cell development in the bone marrow is tightly regulated and as both the pre-BCR and BCR use NF-κB as part of their signaling cascades the role of members of this family of transcription factors has long been of interest.2 To date, effects on bone marrow B-cell development were only observed using approaches simultaneously deleting genes to inhibit both NF-κB pathways10,14,15 or constitutive activation of the classical pathway.13 Earlier developmental stages were affected in several models3,16,21 with ectopic Pax5 expression rescuing B-cell development. Although constitutive activation of either the alternative18 or classical NF-κB pathways12 using CD19 Cre affected mature B cells in the periphery, there was no reported impact on bone marrow development. We observed a similar accumulation of peripheral B cells using mb1 cre (Figure 1), likely because of independence from the pro-survival BAFF signaling described in the CD19 cre models12,18 as mb1 is expressed just slightly earlier in development.1 However, constitutive activation of both pathways simultaneously did not result in further accumulation of B cells, with mice displaying a marked reduction in B cells in all peripheral organs and in the blood (Figure 1).
To confirm that this B-cell deficiency was due to a developmental issue, we looked at the different B-cell populations in the bone marrow and found that not only were B220+ cells markedly reduced (Figure 2), the late pro-B/fraction C cells were almost completely absent (Figure 2), coinciding with when the expression of mb1 first occurs in the entire population of developing B cells (supplemental Figure 4). Deletion or mutation of mb1 results in an ability to express a pre-BCR and in arrest of B-cell development at the pro-B-cell stage.22,23 Because the mb1DPWY mice are heterozygous for mb1 cre, they still express Igα and thus still capable of expressing the pre-BCR.
Another cause of developmental arrest can be due to the lack of surface IgM expression, as observed in μMT mice,24 whereas overexpression of μ heavy chain also results in a blockade in B-cell development at the pro-/pre-B-cell transition.25 Both models result in an accumulation of intracellular IgM; however, splenic B cells in our model express surface IgM at only a modestly reduced level compared with controls (Figure 4 and data not shown) and are functionally able to respond to IgM stimulation (Figure 6), suggesting that this is not the cause of developmental arrest.
Generation of cotransfer chimeras allowed us to demonstrate that the observed defect is not only B cell–intrinsic but also cell autonomous, with WT stroma and B cells unable to rescue B-cell development, a result reinforced by in vitro culture experiments (Figure 3). Analysis of Caspase 3 and 7 found that there were significantly more late Pro-B/fraction C cells undergoing apoptosis (Figure 4B), a similar result was observed in early B cells in IL-7R knockout mice where the early development stages reliant on IL-7 for survival signals are affected.26,27 Analysis of genes related to apoptosis found increased expression of pro-apoptotic genes and decreased expression of anti-apoptotic genes in the early pro-B cell/fraction B population, of which Bim is known to play an important role in pro-B cells28 (supplemental Figure 5). Additionally, FAS and FASL expression were not detected, adding confirmation to this being a cell intrinsic phenomenon. Given the cross regulation between pre-BCR and IL-7 signaling so that developing B cells can switch between VDJ recombination and proliferation,29 it is conceivable that constitutive activation of both NF-κB pathways sufficiently perturbs this balance to result in apoptosis of the majority of B cells.
A key transcription factor in B cell development is Pax5, mice lacking Pax5 lack B cells from early pro-B cells onward and these cells fail to express CD19.30 B cells in mb1DPWY mice have normal CD19 expression. Using quantitative polymerase chain reaction analysis, we found significantly reduced expression of the B cell–obligate transcription factors Ebf131 and Pax530,32 as well as Ikaros,32 which is important for IgH recombination via RAG1/2 expression (Figure 4E). This suggests a significant perturbation of the B-cell transcriptional program, leading to inappropriately developing B cells that are removed by apoptosis as they fail to progress through the hierarchical development pathway.
A further wave of apoptosis is observed in immature B cells/fraction E, suggesting that cells are unable to function appropriately (Figure 4B). Indeed, the small proportion of cells that progress and populate the periphery were largely of an immature phenotype compared with controls suggesting further effects, potentially because of mimicry of tonic BCR signaling. These cells mount an impaired proliferative response to stimulation involving NF-κB signaling such as LPS and CD40 (Figure 6A-B), although surprisingly the proliferative response to BCR signaling appeared to remain intact (Figure 6B) and cells were able to home to secondary lymphoid organs and formed small B cell follicles in the spleen (Figure 5C; supplemental Figure 2). In line with a lack of mature peripheral B cells, mb1DPWY mice were found to have extremely low levels of circulating antibody and lacked the predisposition to autoimmune disease observed in the other strains tested (Figure 6).
Generating CD21DPWY mice, where activation starts in immature B cells, which will egress from the bone marrow, did not have any loss of B cells, and instead had an accumulation of mature cells in the periphery (Figure 5). This was particularly interesting because an original goal of the study was to determine whether early constitutive activation would drive tumor development, as various B-cell malignancies have mutations activating both NF-κB pathways.33,34 Our analysis indicates that these probably result from genetic alterations acquired after B cells have already passed through critical early developmental stages because an earlier activation would likely have resulted in deletion of the cell.
The development of B cells in the bone marrow is a complex process, tight coordination of survival and proliferation signals are coupled with the need for the generation and selection of a functional, non-autoreactive BCR. The complexity of these signals necessitate multiple checkpoints at which cells can be deleted. The data presented here, when taken together with the work of others, demonstrate the integral and highly regulated role that the NF-κB pathways play in this process. Perturbations can lead to the complete loss of B cells at various points in development, and we provide evidence that even signals that would normally be deemed helpful in promoting survival and proliferation early in development as they do in the periphery and late bone marrow development, can instead interfere massively with the earlier developmental process.
Requests for data sharing should be e-mailed to the corresponding author, Andrea Paun (pauna@nih.gov).
Acknowledgments
The authors acknowledge Hongshan Wang for technical assistance and other members of the Siebenlist laboratory for helpful discussions.
This research was supported by the Intramural Research Program of National Institute of Allergy and Infectious Diseases, National Institutes of Health.
Authorship
Contribution: A.P. designed studies, performed experiments, analyzed data, and wrote the manuscript; E.C. designed studies, helped organize animal experiments, and discussed results; and U.K.S. designed studies, analyzed the data, and wrote the manuscript.
Conflict-of-interest disclosure: The authors declare no competing financial interests.
The current affiliation for A.P. is Laboratory of Parasitic Diseases, National Institute of Allergy and Infectious Diseases, National Institutes of Health, Bethesda, MD.
Correspondence: Andrea Paun, Laboratory of Molecular Immunology, National Institute of Allergy and Infectious Diseases, National Institutes of Health, 4 Memorial Dr, Building 4, Room B1-12, Bethesda, MD 20814; e-mail: pauna@nih.gov; and Estefania Claudio, Laboratory of Molecular Immunology, National Institute of Allergy and Infectious Diseases, National Institutes of Health, 50 South Dr, Building 50, Room 6241, Bethesda, MD 20814; e-mail: evazquez@niaid.nih.gov.

