

Abstract
Hematopoiesis in teleost fish is maintained in the kidney. We previously reported that Hoechst dye efflux activity of hematopoietic stem cells (HSCs) is highly conserved in vertebrates, and that Hoechst can be used to purify HSCs from teleost kidneys. Regulatory molecules that are strongly associated with HSC activity may also be conserved in vertebrates. In this study, we identified evolutionarily conserved molecular components in HSCs by comparing the gene expression profiles of zebrafish, murine, and human HSCs. Microarray data of zebrafish kidney side population cells (zSPs) showed that genes involved in cell junction and signal transduction tended to be up-regulated in zSPs, whereas genes involved in DNA replication tended to be down-regulated. These properties of zSPs were similar to those of mammalian HSCs. Overlapping gene expression analysis showed that 40 genes were commonly up-regulated in these 3 HSCs. Some of these genes, such as egr1, gata2, and id1, have been previously implicated in the regulation of HSCs. In situ hybridization in zebrafish kidney revealed that expression domains of egr1, gata2, and id1 overlapped with that of abcg2a, a marker for zSPs. These results suggest that the overlapping genes identified in this study are regulated in HSCs and play important roles in their functions.
Introduction
The hematopoietic system is made up of a complex hierarchy of cells at various stages of differentiation. Hematopoietic stem cells (HSCs) are the most immature elements of the hematopoietic hierarchy, and can produce different classes of hematopoietic progenitor cells (HPCs). Only HSCs are capable of lifelong self-renewal, and are the operative population in hematopoietic transplantation. Understanding how HSC fate is controlled is therefore of critical importance. HSCs are regulated by a complex network of gene interactions that maintain the proper balance between self-renewal and differentiation. Global gene expression profiles of purified HSCs have identified numerous candidate HSC regulatory molecules.1-3 Although some molecules and pathways have been implicated in HSC self-renewal, including HoxB4, Bmi-1, and the Wnt/β-catenin signaling pathway,4-6 the genetic programs regulating HSCs remain poorly understood.
Hematopoiesis in teleost fish is maintained in the kidney. Definitive blood cell lineages in teleost fish show a high degree of similarity at the morphologic and molecular level with their mammalian counterparts.7 The zebrafish (Danio rerio) has proven utility as a genetically tractable vertebrate organism, and many transgenic and mutant zebrafish have been used to study hematopoiesis. The key regulatory genes responsible for generating hematopoietic cells or lineage-committed cells have been analyzed in zebrafish.8 The major blood cell lineages can be isolated from zebrafish kidney by flow cytometry (FCM) based on differences in light scatter profiles. Traver et al showed that HSCs were contained only in the lymphoid population in zebrafish kidney by performing transplantation of hematopoietic cells into gata1−/− mutant embryo.9 However, the resolution of this procedure is not sufficient to purify HSCs from zebrafish kidney. Because methods for purifying HSCs have not been established, gene expression profiles of HSCs have not yet been reported in zebrafish. Recently, we developed a transplantation model system of hematopoietic cells using clonal ginbuna crucian carp (Carassius auratus langsdorfii) and demonstrated that HSCs reside in the kidney.10 We also reported that side population (SP) cells, identified based on their capacity to efflux the fluorescent dye Hoechst 33342 (Hoechst) in an FCM analysis, were enriched for HSCs in teleost kidney,11,12 as has been described in mammalian bone marrow.13 Moreover, we recently showed that Hoechst dye efflux activity of SP cells in zebrafish kidney was attributed to the expression of abcg2a,14 a homolog of ABCG2, which is associated with Hoechst dye efflux activity in HSCs.15 These findings indicate that the Hoechst dye efflux activity of HSCs is highly conserved in vertebrates, and that Hoechst can also be used to purify HSCs from teleost fish.
Several groups have attempted to characterize the molecular signature of stem cells by comparing the gene expression profile of murine HSCs (mHSCs) with the profiles of murine neural or embryonic stem cells.1,2,16 These analyses revealed so-called “stemness” genes, which are proposed to be important for conferring the functional characteristics of stem cells. However, the overlapping genes in these stem cells were extremely limited.16 In contrast, mHSCs and human HSCs (hHSCs) share several expressed gene products.1 Regulatory molecules that are strongly associated with HSC activity may be conserved in phylogenetically more distant species, such as humans and zebrafish. The zebrafish kidney SP cells (zSPs) are particularly suitable for identifying the expression profile of teleost HSCs. In the present study, to identify evolutionarily conserved molecular components in HSCs, we sought to compare the gene expression profile of zSPs with that of murine and human bone marrow HSCs using microarray analysis.
Methods
Fish
Wild-type zebrafish were obtained from a local pet supplier, kept at 28°C in a recirculation system with filtered water, and fed pelleted dry food twice daily. Because hematopoietic cells isolated from male zebrafish kidney were frequently contaminated with testicular cells, and most testicular cells showed low Hoechst fluorescence,14 only female zebrafish were used in the present study to prevent contamination. This study was carried out according to a protocol approved by the institutional review boards of all participating institutions.
Cell preparation and FCM
Hematopoietic cells were obtained from adult zebrafish kidney, as previously described.14 Briefly, trunk (body, mesonephric, posterior) kidney was dissected, and hematopoietic cells were obtained by macerating the kidney on a stainless steel mesh in 5 mL of ice-cold Hanks balanced salt solution (HBSS) containing 2% of fetal bovine serum. Cells from trunk kidney were pelleted by centrifugation. After discarding the supernatant, the pellet was gently mixed with 1 mL of distilled water by pipetting to lyse the erythrocytes by osmotic shock. Subsequently, 1 mL of 2× HBSS was added, and the cells were washed twice with HBSS by centrifugation. Cells were resuspended at a density of 106 cells/mL in HBSS. Hematopoietic cells from 8 to 20 zebrafish were stained with 3 μg/mL Hoechst 33342 (Hoechst; Molecular Probes) for 90 minutes at 25°C. After staining, cells were washed by centrifugation, adjusted to 5 × 106 cells/mL in HBSS, and kept on ice until use. Just before FCM analysis, propidium iodide (Molecular Probes) solution was added at a final concentration of 2 μg/mL to identify nonviable cells. FCM analysis was performed on a dual laser (488 nm and ultraviolet) flow cytometer (EPICS ALTRA; Beckman Coulter), as previously described.11
For cell cycle analysis, cells were stained with 3 μg/mL Hoechst for 45 minutes at 25°C. Then, Pyronin Y (Sigma-Aldrich) was added at a final concentration of 1 μg/mL, and cells were incubated for another 45 minutes.
RNA isolation and microarray processing
To minimize biologic variability, 20 zebrafish were used for 3 independent cell-sorting experiments, and lysates from these 3 experiments were pooled by cell type. Cells were sorted into lysis buffer directly, and total RNAs from 1.6 × 104 cells of each subset (zSPs and zMPs; see Figure 1A-B) were extracted using the Absolutely RNA Nanoprep Kit (Stratagene). To determine the reliability of microarray processing, each RNA sample was split into 2 aliquots and used for amplification, labeling, and hybridization to independent arrays. In the first amplification round, polyinosinic acid (Sigma-Aldrich) was added into the RNA samples as a carrier, and RNAs were amplified using the MessageAmp II aRNA Amplification Kit (Applied Biosystems) for 14 hours at 37°C. Amplified cRNAs were then purified and used for the second round of amplification. In the second amplification round, cRNAs were amplified and labeled with biotin-11-uridine triphosphate (Applied Biosystems) for 14 hours at 37°C. After biotin-labeled cRNAs were purified, they were fragmented and hybridized to the GeneChip Zebrafish Genome Array (Affymetrix). Hybridization, washing, and staining with streptavidin-phycoerythrin conjugate, and scanning were done essentially as recommended by Affymetrix.
Data analysis
After scanning, the normalization of raw data was performed in R (version 2.6.2, http://www.r-project.org/) using the Robust Multichip Array algorithm, which is implemented in the affy package of Bioconductor (http://www.bioconductor.org/). To evaluate the quality and the reproducibility of these data, Pearson correlation coefficients were calculated using Microsoft Excel, and principal components analysis was performed using R. The correlation of microarray data for the 2 replicates of each subset was extremely high (supplemental Figure 1A, available on the Blood website; see the Supplemental Materials link at the top of the online article). The principal components analysis demonstrates that the 2 replicates of each subset cluster closely together (supplemental Figure 1B). These data indicate that analysis of individual hybridizations is highly reproducible for each sample. Up- and down-regulated genes in zSPs were selected from normalized data of each sample using rank products (RP). The RP is statistically rigorous and can be used to provide reliable significance thresholds as a P value to distinguish significantly regulated genes.17 Up- and down-regulated genes were defined with a threshold of P less than .05 using the RankProd package of Bioconductor. Affymetrix raw data of mHSCs, mouse HPCs (mHPCs), hHSCs, and human HPCs (hHPCs) samples were downloaded from the Gene Expression Omnibus (GEO; http://www.ncbi.nlm.nih.gov/geo/) database, and up- and down-regulated genes in HSCs were selected by the same method as used for zSPs. The expression signals were calculated from back-transformed (anti-log2) Robust Multichip Array values, and fold change was calculated by dividing the mean expression signal of zSPs, mHSCs, and hHSCs by that of zMPs, mHPCs, and hHPCs, respectively. For Gene Ontology (GO) term enrichment analysis, the hypergeometric-based tests were carried out in the lists of up- and down-regulated genes using the GOstats package (Version 1.7.4) of Bioconductor with a P value cutoff of .05. To compare expressed genes from the zebrafish, mouse, and human datasets, gene correspondences have been made among these 3 species using the tool provided by Affymetrix (NetAffx: http://www.affymetrix.com/jp/analysis/index.affx) and BioMart (http://www.biomart.org/). Genes present on the zebrafish, mouse, and human microarrays were intersected by Entrez gene identification number to determine overlaps (supplemental Figure 2). The hypergeometric distribution was used to obtain the chance probability of observing overlaps. The probability of observing an overlap (x) is given by:
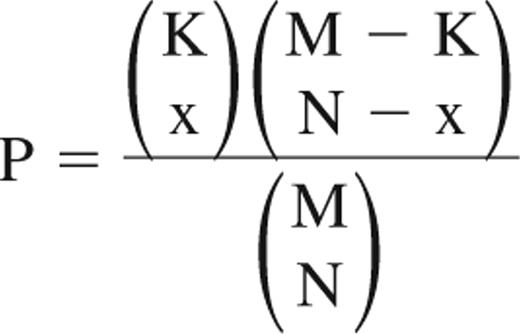
where (K) is the total number of homologous genes within the up- or down-regulated genes in zSPs; (M) is the total number of homologous pairs; and (N) is the number of homologous genes within the up- or down-regulated genes in mHSCs or hHSCs.
The microarray data from zebrafish have been deposited in the GEO database, and have been assigned the following accession numbers: GSM324903, GSM324904, GSM324905, and GSM324906 (Series GSE12955). The GEO accession numbers of microarray data used in this study (mouse and human) are as follows: mouse samples, GSM175730, GSM175731, GSM175734, and GSM175735 (Series GSE7302); human samples, GSM51401, GSM51402, GSM51403, GSM51404, GSM51405, GSM51406, GSM51408, GSM51419, GSM51420, GSM51421, GSM51422, GSM51423, GSM51424, and GSM51426 (Series GSE2666).
Reverse transcription–polymerase chain reaction
For expression analysis, sorted cells were pooled by cell type in lysis buffer, and total RNAs from 5 × 103 cells of each subset were extracted and amplified, as described in the previous paragraph. Amplified RNAs were reverse transcribed into DNA using TaqMan reverse-transcription reagents with random hexamers (Applied Biosystems). Expression analysis by semiquantitative reverse transcription–polymerase chain reaction (RT-PCR) with specific primer sets was performed using BIOTAQ (Bioline), as described previously.14 The primer sequences and Entrez gene identification numbers are listed in supplemental Table 1.
A quantitative real-time polymerase chain reaction (qPCR) assay was performed on an ABI 7500 Fast Real-Time PCR System using TaqMan Fast Universal PCR master mixture (Applied Biosystems). Probes and primers used in qPCR assay are listed in supplemental Table 1. Data were analyzed by 7500 Fast System SDS Software 1.3.1.
In situ hybridization in zebrafish kidney
Digoxigenin (DIG) and fluorescein-labeled RNA probes for egr1, f11r, gata2, id1, krt18, cd45, and abcg2a genes were prepared, as described previously.14 Whole-mount in situ hybridization was carried out in zebrafish kidney following the protocol by Jowett,18 with some modifications. Briefly, trunk kidneys from adult zebrafish were fixed in 4% paraformaldehyde in phosphate-buffered saline (PBS). Before hybridization, kidneys were incubated in 6% H2O2 in PBS and 0.1% Tween 20 for 1 hour to quench endogenous peroxidase activity, and then treated with proteinase K (20 μg/mL, 30 minutes). Hybridization was performed using DIG-labeled antisense RNA probes for 18 hours at 65°C. For detection of DIG-labeled RNA probes, kidneys were blocked in 1× blocking reagent (Roche Molecular Biochemicals), and incubated overnight at 4°C with the alkaline phosphatase-labeled anti-DIG antibody (Roche; 1/2000 dilution). The color reaction was developed using nitroblue tetrazolium chloride and 5-bromo-4-chloro-3-indolyl-phosphate (Roche). Kidneys were then embedded in paraffin, and 4-μm–thick sections were prepared. Visible light imaging was performed on Olympus IX70 microscope using ×40 and ×100 oil objective lens and Olympus DP70 digital camera and software (Olympus).
Double-fluorescent whole-mount in situ hybridization was performed according to the protocol by Sumanas et al,19 with some modifications. Briefly, fixed kidneys were hybridized with both fluorescein-labeled RNA probe (egr1, f11r, gata2, id1, or krt18) and DIG-labeled abcg2a probe. Kidneys were then blocked in 1× blocking reagent, incubated with the horseradish peroxidase (HRP)–labeled anti-fluorescein antibody (Vector Laboratories; 1/500 dilution), and incubated with fluorescein tyramide (PerkinElmer Life Sciences) to amplify fluorescein deposition. To inactivate the HRP activity associated with the detection of the fluorescein-labeled probe, kidneys were incubated in 1% H2O2 in PBS and 0.1% Tween 20 for 30 minutes. Kidneys were then blocked, incubated with the HRP-labeled anti-DIG antibody (Roche Molecular Biochemicals; 1/100 dilution), and incubated with cyanine 3 tyramide (PerkinElmer Life Sciences). Frozen sections of 20 μm in thickness were prepared on a cryostat (Microm HM 550). The images were captured using a confocal laser-scanning microscope (Fluoview FV1000; Olympus) with a ×40 oil objective lens and FV10-ASW (version 1.6a) software (Olympus).
Results
Experimental design and microarray analysis
We previously characterized SP cells and main population (MP, non-SP) cells in zebrafish and ginbuna kidney. Kidney SP cells had a thin-layered cytoplasm and round nucleus,11,14 which are typical morphologic features of HSCs. FCM analysis showed that SP cells were mainly present in the lymphoid population (forward scatterlow; side scatterlow), and that SP cells hardly contained mature lymphocytes, such as CD4-, CD8α-, and immunoglobulin M–positive cells, although MP cells in the lymphoid population contained several mature lymphocytes.12 Transplantation experiments using clonal ginbuna showed that kidney SP cells contained a high number of HSCs, but a low number of HPCs. In contrast, MP cells in the lymphoid population contained a high number of HPCs.11,12 In the present study, to identify regulated genes in zebrafish HSCs, lymphoid cells from zebrafish kidney were subdivided into the SP and MP populations (referred to as zSPs and zMPs, respectively), and gene expression analysis was performed on each cell population using a microarray (Figure 1A-B). Up- and down-regulated genes in zSPs were selected by comparison analysis of zSPs to zMPs using RP. As shown in Figure 1C, we identified 1077 (9.8%) and 913 (8.3%) up- and down-regulated genes from a total of 10 971 genes, respectively. The top 80 statistically significant up- and down-regulated genes in zSPs are listed in supplemental Tables 2 and 3, respectively.
![Figure 1. Identification of up- and down-regulated genes in zSPs. Hematopoietic cells from zebrafish kidney were stained with Hoechst 33342 (Hoechst) and analyzed by FCM. (A) Scatter profile of kidney hematopoietic cells. Gated region indicates lymphoid cells (forward scatter [FS]low; side scatter [SS]low). (B) Hoechst fluorescence of the lymphoid cells. The lymphoid cells were subdivided into 2 populations, zSPs and zMPs. (C) The numbers of up- and down-regulated genes of zSPs. A total of 10 971 genes is present on the GeneChip Zebrafish Genome Array. Among them, 1077, 8981, and 913 genes were up-regulated, unchanged, and down-regulated in zSPs, respectively.](/view-large/figure/8162085/zh89990946700001.jpeg)
Identification of up- and down-regulated genes in zSPs. Hematopoietic cells from zebrafish kidney were stained with Hoechst 33342 (Hoechst) and analyzed by FCM. (A) Scatter profile of kidney hematopoietic cells. Gated region indicates lymphoid cells (forward scatter [FS]low; side scatter [SS]low). (B) Hoechst fluorescence of the lymphoid cells. The lymphoid cells were subdivided into 2 populations, zSPs and zMPs. (C) The numbers of up- and down-regulated genes of zSPs. A total of 10 971 genes is present on the GeneChip Zebrafish Genome Array. Among them, 1077, 8981, and 913 genes were up-regulated, unchanged, and down-regulated in zSPs, respectively.
Identification of up- and down-regulated genes in zSPs. Hematopoietic cells from zebrafish kidney were stained with Hoechst 33342 (Hoechst) and analyzed by FCM. (A) Scatter profile of kidney hematopoietic cells. Gated region indicates lymphoid cells (forward scatter [FS]low; side scatter [SS]low). (B) Hoechst fluorescence of the lymphoid cells. The lymphoid cells were subdivided into 2 populations, zSPs and zMPs. (C) The numbers of up- and down-regulated genes of zSPs. A total of 10 971 genes is present on the GeneChip Zebrafish Genome Array. Among them, 1077, 8981, and 913 genes were up-regulated, unchanged, and down-regulated in zSPs, respectively.
To compare the regulated genes in zSPs with those in mammalian HSCs, published datasets of HSCs and HPCs in murine and human bone marrow were obtained from the GEO database. Previous reports have shown that mHSCs in bone marrow are enriched in the lineage-negative (Lin−) Sca-1+ c-kit+ CD34− Flt3− population, whereas HPCs are enriched in the Lin− Sca-1+ c-kit+ CD34+ Flt3+ population.20,21 In the present study, therefore, microarray data of these phenotypic mHSCs and mHPCs were used for comparative gene expression analysis. In contrast, hHSCs and hHPCs in bone marrow are enriched in the CD34+ CD33− CD38− c-kit+ rhodamine-123low (rholow) population and the CD34+ CD33− CD38− rhohigh population, respectively,22 and microarray data of these phenotypic hHSCs and hHPCs were used. Up- and down-regulated genes in mHSCs and hHSCs were also selected by comparison analysis of HSCs to HPCs using RP (supplemental Figure 3).
GO term enrichment analysis
GO is a gene classification tool that is used to categorize and understand the biologic role of genes. A large number of genes are classified based on the 3 GO categories, cellular component, molecular function, and biological process. GO term enrichment analysis reveals which GO terms are significantly overrepresented in the differentially expressed genes.23 To examine the biologic properties of the up- and down-regulated genes of zSPs, GO term enrichment analysis was performed with each gene list. Genes involved in cell junction, extracellular matrix, intracellular signaling cascade, and signal transduction were overrepresented in the up-regulated genes of zSPs (Table 1). This suggests that SP cells in zebrafish kidney adhere to their microenvironment and express many kinds of signaling molecules. In contrast, genes involved in translation, RNA processing, DNA repair, and DNA replication were overrepresented in the down-regulated genes of zSPs (Table 2), suggesting that SP cells are in a quiescent state of the cell cycle and have a low potential for posttranscriptional regulation. To confirm that SP cells are quiescent, we examined the cell-cycle status of zSPs and zMPs by Pyronin Y staining. The percentage of G0 cells in zSPs (85.0%) was significantly higher than that in zMPs (57.8%), indicating that SP cells in zebrafish kidney are relatively quiescent compared with MP cells (supplemental Figure 4).
Because the GO vocabulary is not specific to any species, this method allows for cross-species comparisons of gene lists. We also performed GO term enrichment analysis with the up- and down-regulated genes of mHSCs and hHSCs. Genes involved in cell junction or cell-cell adherens junction and intracellular signaling cascade were overrepresented in the up-regulated genes of mHSCs and hHSCs (supplemental Tables 4 and 6). In contrast, genes involved in DNA repair and DNA replication were overrepresented in the down-regulated genes of mHSCs and hHSCs (supplemental Tables 5 and 7). These results of mHSCs and hHSCs were similar to those of zSPs, suggesting that zSPs possess the general molecular hallmarks of HSCs, and that HSCs from these 3 species share a core set of expressed gene products.
Overlapping gene expression analysis of HSCs
Conducting a comparison of the gene expression profile of zSPs with that of mHSCs and hHSCs would be a useful strategy for identifying expression signatures of HSC activity. To identify evolutionarily conserved molecular components in HSCs, we compared the up- and down-regulated genes of zSPs with those of mHSCs and hHSCs. The data show that several genes are commonly up- or down-regulated in these 3 types of HSCs (Figure 2). The up-regulated genes of zSPs (1077 genes) overlapped with 168 (15.6%) and 131 (12.1%) genes up-regulated in mHSCs and hHSCs, respectively, and 40 (3.7%) genes were commonly up-regulated in these 3 HSCs (Figure 2A). These 40 homologous genes are listed in Table 3. Genes up-regulated in all 3 HSCs belong to a variety of GO terms, including transcription factor activity (eg, egr1, gata2, and id1), cell junction (eg, dst, f11r, and perp), and extracellular space (eg, apoe, tfpi, and timp2). In contrast, the down-regulated genes of zSPs (913 genes) overlapped in 136 (14.9%) and 172 (18.8%) genes with those of mHSCs and hHSCs, respectively, and 47 (5.1%) genes were commonly down-regulated in these 3 HSCs (Figure 2B). These 47 genes include translation-related genes, such as eif1ay, and DNA replication-related genes, such as pola1, pole2, and rfc4 (supplemental Table 8).

Overlapping sets of expressed genes in zebrafish HSCs, mHSCs, and hHSCs. Datasets of zebrafish (zSPs and zMPs), mice (mHSCs and mHPCs), and humans (hHSCs and hHPCs) were analyzed with the same method. Venn diagrams of the number of up-regulated (A) and down-regulated genes (B) are shown. Forty and 47 genes were commonly up- and down-regulated in these 3 HSCs, respectively.
Overlapping sets of expressed genes in zebrafish HSCs, mHSCs, and hHSCs. Datasets of zebrafish (zSPs and zMPs), mice (mHSCs and mHPCs), and humans (hHSCs and hHPCs) were analyzed with the same method. Venn diagrams of the number of up-regulated (A) and down-regulated genes (B) are shown. Forty and 47 genes were commonly up- and down-regulated in these 3 HSCs, respectively.
To evaluate the significance of zSPs in relation to mHSCs and hHSCs, we calculated the probability of observing such an overlap by chance using the hypergeometrical distribution (supplemental Table 9). We identified 774 mouse homologs and 768 human homologs of zSPs up-regulated genes (1077 genes). Of these, 168 (21.7%) were up-regulated in mHSCs and 131 (17.1%) were up-regulated in hHSCs. The probabilities of these overlaps occurring by chance were extremely low (P < .001). Similarly, the probabilities of the overlaps occurring in down-regulated genes by chance were extremely low. Furthermore, the percentage of mouse homologs for hHSCs-up-regulated genes that were up-regulated in mHSCs (22.3%; hHSCs vs mHSCs) was very similar to that between zSPs and mHSCs (21.7%). These results indicate that the expression profiles of HSCs are highly conserved among vertebrates, and that these overlapping genes represent the conserved molecular components in vertebrate HSCs.
Validation of microarray results by RT-PCR and in situ hybridization
To confirm that the up- and down-regulated genes identified from microarray analysis were actually regulated in HSCs, we carried out semiquantitative RT-PCR analysis with RNA samples derived from zSPs and zMPs. The expression levels of 8 genes (apoe, egr1, f11r, gata2, id1, krt18, meis1, and timp2) in zSPs, which were identified as common up-regulated genes in vertebrate HSCs (Table 3), were higher than those in zMPs (Figure 3A). In contrast, the expression levels of 4 genes (eif1ay, h2afz, pola1, and rfc4) in zSPs, which were identified as common down-regulated genes (supplemental Table 8), were lower than those in zMPs (Figure 3B). We further confirmed expression levels of 5 up-regulated genes (egr1, f11r, gata2, id1, and krt18) and 2 down-regulated genes (eif1ay and rfc4) in zSPs and zMPs by qPCR (Figure 4D-E). In no case did semiquantitative RT-PCR or qPCR analysis contradict the microarray data. These results indicate that the microarray analysis of zSPs and zMPs is able to detect the differentially expressed genes in a reproducible manner.

Expression analysis of up- and down-regulated genes in zSPs and zMPs. (A-C) Semiquantitative RT-PCR was performed in zSPs and zMPs. The expression levels of up-regulated genes (apoe, egr1, f11r, gata2, id1, krt18, meis1, and timp2) (A) and down-regulated genes (eif1ay, h2afz, pola1, and rfc4) (B) were compared between zSPs and zMPs. cd45 and ef1α were used as a blood cell marker and an internal control, respectively (C). NC indicates negative control. (D-E) A qPCR assay was performed in zSPs, zMPs, and zebrafish kidney. Expression of the ef1α gene was used to normalize the amount of the investigated transcripts. Relative expression levels in zSPs and zMPs were calculated from the expression levels in zebrafish kidney. Results are presented as an average expression level ± SD (n = 4). There are significant differences between zSPs and zMPs (*P < .001).
Expression analysis of up- and down-regulated genes in zSPs and zMPs. (A-C) Semiquantitative RT-PCR was performed in zSPs and zMPs. The expression levels of up-regulated genes (apoe, egr1, f11r, gata2, id1, krt18, meis1, and timp2) (A) and down-regulated genes (eif1ay, h2afz, pola1, and rfc4) (B) were compared between zSPs and zMPs. cd45 and ef1α were used as a blood cell marker and an internal control, respectively (C). NC indicates negative control. (D-E) A qPCR assay was performed in zSPs, zMPs, and zebrafish kidney. Expression of the ef1α gene was used to normalize the amount of the investigated transcripts. Relative expression levels in zSPs and zMPs were calculated from the expression levels in zebrafish kidney. Results are presented as an average expression level ± SD (n = 4). There are significant differences between zSPs and zMPs (*P < .001).

In situ hybridization in zebrafish kidney. Whole-mount in situ hybridization was carried out in zebrafish kidney. (A-E) The expression signals from egr1 (A), gata2 (B), id1 (C), f11r (D), and krt18 (E) were visualized by nitroblue tetrazolium chloride and 5-bromo-4-chloro-3-indolyl-phosphate. Cells expressing egr1, f11r, gata2, id1, and krt18 mRNA are present on the surface of renal tubules (arrowheads). (F-J) Double-fluorescent whole-mount in situ hybridization was performed. The expressions of egr1, gata2, id1, f11r, and krt18 (left panels) and abcg2a (middle panels) were visualized by fluorescein and cyanine 3, respectively. Right panels show merged images. Cells coexpressing egr1, gata2, id1, f11r, or krt18 and abcg2a are present on the surface of renal tubules (arrowheads). Dotted lines indicate the surface of the renal tubule. All scale bars indicate 5 μm.
In situ hybridization in zebrafish kidney. Whole-mount in situ hybridization was carried out in zebrafish kidney. (A-E) The expression signals from egr1 (A), gata2 (B), id1 (C), f11r (D), and krt18 (E) were visualized by nitroblue tetrazolium chloride and 5-bromo-4-chloro-3-indolyl-phosphate. Cells expressing egr1, f11r, gata2, id1, and krt18 mRNA are present on the surface of renal tubules (arrowheads). (F-J) Double-fluorescent whole-mount in situ hybridization was performed. The expressions of egr1, gata2, id1, f11r, and krt18 (left panels) and abcg2a (middle panels) were visualized by fluorescein and cyanine 3, respectively. Right panels show merged images. Cells coexpressing egr1, gata2, id1, f11r, or krt18 and abcg2a are present on the surface of renal tubules (arrowheads). Dotted lines indicate the surface of the renal tubule. All scale bars indicate 5 μm.
We examined the expression patterns of some common up-regulated genes in zebrafish kidney by in situ hybridization. We focused our analyses on the following genes: (1) egr1 (early growth response 1): Egr1 is a member of the immediate early response transcription factor family. Min et al24 showed that Egr1 expression was enriched in HSCs. However, Egr1 was greatly reduced when HSCs were stimulated to proliferate and migrate. Egr1 deficiency in mice leads to a significant increase in HSC proliferation in the bone marrow, implying that Egr1 target genes promote the retention of HSCs in their niches. (2) gata2 (GATA-binding protein 2): Gata2 is a zinc-finger transcription factor that is most abundant in immature progenitors and plays a role in both the proliferation and survival of early hematopoietic cells.25 Gata2-null embryonic stem cells did not contribute to bone marrow hematopoiesis, indicating the essential roles of Gata2 in the production and maintenance of HSCs.26 (3) id1 (inhibitor of DNA binding 1): Id1 acts as a dominant-negative regulator of basic helix-loop-helix transcription factors. The down-regulation of Id1 is necessary for differentiation to proceed in many cell lineages.27-29 Id1-deficient HSCs show increased cell cycling, but fail to efficiently self-renew.30 (4) f11r (F11 receptor; also known as junctional adhesion molecule-A): F11r is a cell adhesion molecule with 2 immunoglobulin-like domains and is localized at a tight junction of epithelial, endothelial, and hematopoietic cells.31-34 Although its role in hematopoiesis is unclear, F11r is known to play a role in the adhesion of blood cells to endothelial cells.35 (5) krt18 (keratin 18): The Krt18 gene codes for a type I keratin (cytokeratin 18) intermediate filament protein. Cytokeratin 18 expression is a feature of terminally differentiated epithelial cells, including renal tubules.36 However, the role of Krt18 in hematopoiesis is still unknown.
We previously reported that 68% of abcg2a-positive cells in the zebrafish kidney, which are equivalent to SP cells, were localized on the surface of renal tubules and tightly adhered to renal tubule epithelial cells.14 This observation suggests that renal tubule epithelial cells in teleost fish are a key component of the HSC niche, a microenvironment supporting the maintenance of HSCs. Typical results of in situ hybridization for egr1, gata2, id1, f11r, and krt18 mRNA in zebrafish kidney were shown in Figure 4. We detected egr1, gata2, id1, and f11r mRNA only in interstitial blood cells. Among them, 72% (36 of 50), 56% (28 of 50), 68% (34 of 50), and 52% (26 of 50) of egr1-, gata2-, id1-, and f11r-positive cells were present on the surface of renal tubules, respectively (Figure 4A-D). Although krt18 mRNA was detected in both blood cells and renal tubules, 62% (31 of 50) of krt18-positive blood cells were present on the surface of renal tubules (Figure 4E). By contrast, cd45-positive cells, which contain various types of blood cells, including zSPs and zMPs (Figure 3C), were detected only in interstitial tissue, and less than 10% of cd45-positive cells were present on the surface of renal tubules (supplemental Figure 5A). These results indicate that cells expressing these common up-regulated genes are largely localized on the surface of renal tubules.
To test whether SP cells express these common up-regulated genes in kidney tissue, the overlapping expression domains of these up-regulated genes and abcg2a were examined by double-fluorescent in situ hybridization. Double-staining analysis revealed that the majority of abcg2a-positive cells (50%-80%) expressed egr1, gata2, id1, f11r, and krt18 mRNA, whereas abcg2a single-positive cells were rarely observed. We found that most of the double-positive cells were present on the surface of renal tubules (Figure 4F-J). These results indicate that the common up-regulated genes are actually regulated in HSCs in zebrafish kidney.
Discussion
The present study demonstrates the potential of applying comparative gene expression analysis of phylogenetically distant species to the identification of evolutionarily conserved molecular components in HSCs. This analysis allowed us to find expression signatures that have a strong association with HSC activity. The molecules identified in this study most likely play important roles in early hematopoietic decisions, including self-renewal and differentiation into the more proliferative HPCs.
Because HSC activity is the result of multiple orchestrated gene interactions, manipulation of HSC activity is most likely achieved by manipulating transcription factors. Our data showed that the 40 genes were commonly up-regulated in zSPs, mHSCs, and hHSCs (Table 3). Among them, 10 genes, Dach1 (dachshund 1), Egr1, Etv5 (ets variant gene 5), Fosl2 (fos-like antigen 2), Gata2, Id1, Maff (v-maf musculoaponeurotic fibrosarcoma oncogene homolog f), Meis1 (myeloid ecotropic viral integration 1), Per2 (period homolog 2), and Zbtb16 (zinc finger and BTB domain containing 16), encode transcription factors. Five transcription factors identified in this study, namely Egr1, Gata2, Id1, Meis1, and Zbtb16, have been previously implicated in the regulation of HSCs in mammalian bone marrow.24,25,30,37,38 Thus, transcription factors shared by vertebrate HSCs play crucial roles in HSC functions. Although little is known about the roles of Dach1, Etv5, Fosl2, Maff, and Per2 in HSC functions, these genes may also be involved in the regulatory network of HSC self-renewal and/or differentiation.
In murine studies, a subpopulation of osteoblasts is reported to be a definitive regulatory component of the HSC niche.39,40 Key features of HSCs in the niches are that they are quiescent, adhere to surrounding cells, and show an SP phenotype.41 In the present study, in situ hybridization in zebrafish kidney showed that cells expressing common up-regulated genes, such as egr1, gata2, id1, f11r, and krt18, were predominantly localized on the surface of renal tubules (Figure 4A-E). In addition, high numbers of abcg2a-positive cells expressed these common up-regulated genes, and were present on the surface of renal tubules (Figure 4F-J). These results strongly support the hypothesis that renal tubule epithelial cells are a key component of the HSC niche in the teleost kidney and provide an adhesive attachment for HSCs. Indeed, zSPs showed up-regulation of some genes encoding a cell adhesion molecule, such as cdh1 (cadherin 1, E-cadherin), cdh17 (cadherin 17, LI-cadherin), alcam (activated leukocyte cell adhesion molecule), f11r, and some claudin family genes (supplemental Table 2). Sugano et al showed that F11r was strongly expressed in the enriched HSC fraction in murine bone marrow, fetal liver, and aorta-gonad-mesonephros, and that F11r is an excellent marker for isolating HSCs.34 These observations suggest that F11r plays important roles in the adhesion and interaction between HSCs and their niches.
HSCs in mammalian bone marrow are generally in a quiescent state of the cell cycle, and proliferate slowly, whereas HPCs actively proliferate and differentiate. Quiescence itself contributes to the unresponsiveness of HSCs, because differentiation requires proliferation.42 HSCs balance quiescence and cell division in the HSC niche and also maintain the potential for long-term hematopoiesis.40 Similar to HSCs, stem cells for epidermis and hair follicles were identified in the bulge region as slow-cycling cells through a 5-bromodeoxyuridine–labeling experiment.43 It is likely that common basic mechanisms contribute to the regulation of the cell cycle in stem cells. The helix-loop-helix protein Id1 has been implicated in the maintenance of HSCs via its role in the cell cycle regulation.30 Interestingly, the expression of the Id1 gene is regulated in part by Egr1,44 and the expressions of the Id1 and Egr1 mRNA in HSCs are conserved among vertebrates (Table 3). These observations suggest that Id1 and Egr1 play a role in HSC-fate decisions by acting as inhibitors of cell cycle progression and differentiation.
A major breakthrough in zebrafish research came with forward genetic mutagenesis screens. Many novel genes that are important in hematopoiesis have been positionally cloned using blood cell mutants, and their biochemical pathways have been investigated. The expression of gata1 and gata2 marks the emergence of the first hematopoietic cells in the embryo. Several blood mutant fish, such as bloodless, spadetail, and cloche, showed changes in gata1 and/or gata2 expression patterns, providing how these genes are activated during embryogenesis.45 Thus, the zebrafish model provides several unique experimental approaches for understanding the function and regulation of genes in hematopoiesis. However, these studies are extremely limited in hematopoietic development, and little is known about the mechanisms of HSC self-renewal and differentiation in zebrafish. The microarray data of zSPs, combined with mutagenesis approaches, may greatly facilitate genetic analysis of HSCs in zebrafish.
Comparative gene expression analysis has been used in cancer research. Gene expression profiles reflecting similar cancer phenotypes are usually conserved between mice and humans.46,47 Microarray data revealed that many similar genes are expressed in liver tumors from zebrafish and humans, which raises the possibility of using zebrafish to model human cancer.48 The present results reveal molecular similarities between zebrafish and mammalian HSCs, and demonstrate that zSPs, mHSCs, and hHSCs share several expressed gene products. Because the zebrafish genome sequence has not been completed, and the number of genes present on the zebrafish microarray is only 11 000, some genes with important roles in hematopoiesis may have been untested. As the zebrafish genome sequences will be completely determined in the near future, additional overlapping genes in zebrafish and mammalian HSCs will be identified. In addition, gene expression profiles of SP cells from other vertebrate species (eg, Xenopus tropicalis) may be useful for identifying the expression signature of HSCs, because the Hoechst dye efflux activity is highly conserved in vertebrates.11,13,14 Further research using zebrafish and other vertebrate species may eventually identify the genetic mechanisms involved in HSC self-renewal and differentiation.
The online version of this article contains a data supplement.
The publication costs of this article were defrayed in part by page charge payment. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734.
Acknowledgment
This work was supported in part by a Grant-in-Aid for Encouragement of Young Scientists from the Japan Society for the Promotion of Science.
Authorship
Contribution: I.K. designed the research, performed experiments, analyzed data, and wrote the paper; H.O. analyzed data; and T.M., K.K., T.N., and T.S. designed the research and edited the manuscript.
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Correspondence: Isao Kobayashi, Department of Cell Differentiation, Sakaguchi Laboratory, School of Medicine, Keio University, 35 Shinanomachi, Shinjuku-ku, Tokyo, Japan; e-mail: kobaisao@sc.itc.keio.ac.jp.

