

In this issue of Blood, demonstrated that activated protein C (APC) reduced the severity of pulmonary complications in a murine model of chronic graft-versus-host disease (cGVHD). The protective effect of APC against cGVHD was mediated by a biased signaling of protease-activated receptor 1 (PAR1) on T lymphocytes.1
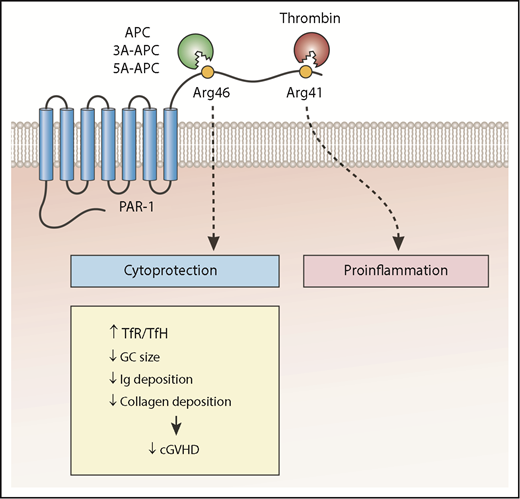
The cytoprotective effect of PAR1 non-canonical signaling after cleavage of PAR1 by wild-type APC or mutant APC (3A-APC or 5A-APC) reduces the number of TFH cells relative to TFR cells, the size of GCs, and antibody and collagen deposition in lungs, which improves bronchiolitis obliterans in a murine model of cGVHD. Ig, immunoglobulin.
The cytoprotective effect of PAR1 non-canonical signaling after cleavage of PAR1 by wild-type APC or mutant APC (3A-APC or 5A-APC) reduces the number of TFH cells relative to TFR cells, the size of GCs, and antibody and collagen deposition in lungs, which improves bronchiolitis obliterans in a murine model of cGVHD. Ig, immunoglobulin.
cGVHD causes morbidity and mortality after allogeneic hematopoietic stem cell transplantation (allo-HSCT) and occurs with various degrees of severity in 30% to 70% of recipients.2 In addition to the organs usually targeted by acute GVHD (aGVHD) (skin, liver, and gastrointestinal tract), cGVHD can affect lungs, mouth, esophagus, joints, muscles, fasciae, eyes, hair, nails, and genitalia. Fibrotic changes in the lungs can result in bronchiolitis obliterans (BO), which is characterized by thickening of the bronchial wall, narrowing of the bronchial lumen, and trapping of air. The pathophysiology of cGVHD involves an early inflammatory phase induced by tissue injury from aGVHD, cytotoxic drugs, and infections. The early inflammation is mediated by innate immunity, which initiates an adaptive immune response that includes B lymphocyte hyper-responsiveness and Th17 differentiation of CD4+ T lymphocytes. This early phase is followed by chronic inflammation and dysregulation of adaptive immunity. Thymic lesions disrupt negative selection of T cells (central tolerance). Reduction in regulatory T (Treg) cells, T follicular regulatory (TFR) cells, regulatory B cells, and regulatory natural killer cells diminish peripheral tolerance. Expansion of B cells in germinal centers (GCs) assisted by T follicular helper (TFH) cells results in the production of pathogenic auto- or alloantibodies. The final phase of cGVHD is associated with fibrosis in various organs.2
GVHD, particularly steroid-refractory GVHD, is associated with endothelial loss of thrombomodulin (TM).3 Preclinical studies showed that soluble TM reduces the severity of GVHD.4 Thrombin binds to TM and activates protein C bound to the endothelial protein C receptor. APC acts as a natural anticoagulant by cleaving activated factor V and factor VIII. However, APC can also cleave protease-activated receptors (PARs)5 (see figure). Cleavage of PARs by APC does not affect the coagulation cascade, but rather results in cytoprotective effects.5 Recombinant mutant APC molecules that are unable to bind factor Va (by replacing lysine residues 191, 192, and 193 with alanine [3K3A-APC or 3A-APC], replacing arginine residues 229 and 230 with alanine, or both [5A-APC]) are not anticoagulants, but they retain their cytoprotective effects6 and have been used in various preclinical sepsis or stroke models and in the case of 3A-APC in early clinical trials.6
APC is not the only protease that cleaves PAR1; thrombin cleaves PAR1, although at a different amino acid residue. APC cleaves PAR1 at arginine residue 46 (Arg46), but thrombin cleaves PAR1 at Arg41 (see figure). Cleavage at 2 different sites results in different post-PAR1 signaling events (biased signaling) and biological function.5,7 Thrombin and thrombin-generated tethered ligand activate G protein signaling and downstream RhoA, resulting in proinflammatory effects. APC and APC-generated tethered ligand (TR47) signals through β-arrestin-2 and Rac1 and results in antiapoptotic and anti-inflammatory effects.5
In a previous study, Ranjan et al3 showed that APC protected against aGVHD in a murine model. The protective effect of APC against aGVHD was mediated by cleavage of PAR3 on T lymphocytes. Cleaved PAR3 together with PAR2 promoted Treg differentiation and Treg expansion, which in turn reduced alloreactive T cells. Pre-incubation of T cells with APC also reduced alloreactivity in vitro and in GVHD in a murine model of bone marrow transplantation.
The authors in the study by Sinha et al showed that daily administration of APC, 3A-APC, or 5A-APC starting 4 weeks after transplantation and continuing for 4 weeks improved pulmonary function tests (reduced airway resistance and pulmonary elasticity and increased compliance) in a murine model of cGVHD. In additional experiments, the authors demonstrated that the protective effect of 5A-APC was dependent on the cleavage of PAR1 on donor T cells. Infusion of donor T cells with mutated PAR1 at the cleavage site of APC (Arg46Gln) removed the protective effect of 5A-APC against BO. Dysregulation of adaptive immunity characterized by a reduction in TFR cells relative to TFH cells and expansion of GCs containing auto- and alloantibody-producing B cells in lymphoid organs are among the pathophysiologic features of cGVHD in mice.2,8-10 Administration of APC, 3A-APC, or 5A-APC increased TFR relative to TFH, reduced antibody and collagen deposition in the lungs, and improved BO.
The Sinha et al study expands the potential therapeutic benefits of APC to cGVHD. The fact that the protective effect of APC against BO is independent of its anticoagulant effect raises the intriguing possibility of using mutant APC molecules (3A-APC and 5A-APC) or their tethered ligand (TR47) in allo-HSCT. However, additional mechanistic data and preclinical studies to investigate the impact of APC on the sclerotic changes in other organs and on the graft-versus-leukemia effect are required before embarking on clinical studies.
Conflict-of-interest disclosure: The author declares no competing financial interests.

This feature is available to Subscribers Only
Sign In or Create an Account Close Modal