

Previous studies by us and others have linked germline genetic variants to the familial predisposition to childhood B-cell acute lymphoblastic leukemia (B-ALL), with pathogenic variants discovered in TP53, PAX5, ETV6, and IKZF1 (J. Clin. Oncol 2018, Nature Genet 2014, Lancet Oncol 2015, Cancer Cell 2018). However, genetic predisposition to T-ALL is much less understood. Rare care reports of T-ALL pedigrees with germline RUNX1 point to its potential role in ALL susceptibility. RUNX1 plays significant roles in definitive hematopoiesis and primarily functions as a transcription factor. RUNX1 germline variants are associated with familial platelet disorder, with a significant proportion of patients also developing myeloid malignancies.
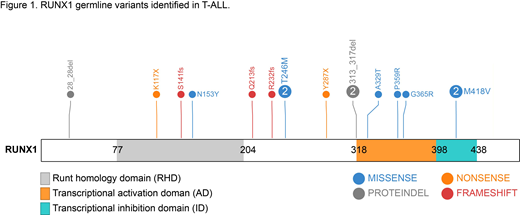
To comprehensively examine the pattern and prevalence of RUNX1 germline variation in T-ALL, we performed targeted germline sequencing of 1,231 cases enrolled on the Children's Oncology Group AALL0434 trial. In this largely unbiased T-ALL cohort, we identified 13 germline RUNX1 variants in 16 cases (Figure 1), including six missense (46.2%), two nonsense (15.4%), three frameshift (23.1%), and two indel variants (15.4%). These variants are divided into three groups: Group I, truncating both the DNA-binding RHD domain and the transcriptional activation AD domain (p.K117* and p.S141fs); Group II, truncating the AD domain only (p.Q213fs, p.R232fs, and p.Y287*); and Group III, missense and indel variants.
To comprehensively characterize the function of these T-ALL-related RUNX1 variants, we performed a variety of biochemical and cellular assays in different model systems. Using reporter gene assays, we first directly evaluated the transcriptional activity of RUNX1 variants in Hela cells and identified both loss-of-function (e.g., Group I variants) and dominant-negative effects (e.g., p.G365R in Group III variants). Group I variants also showed dramatic subcellular mislocalization in the cytoplasm, with concomitant loss of CBFβ binding, both of which were significantly subtler for Groups II and III variants. Focusing on representative variants in these three groups (p.S141fs, p.R232fs, Y287*, and p.G365R), we next examined their effects on hematopoietic phenotypes in vitro. Ectopic expression of Group II and III variants in human CD34+ cells significantly increased CFU-M/GM colony formation and long-term proliferation, while repressing BFU-E colonies. Variant RUNX1 cells also showed defects in megakaryocyte and pre-T cell differentiation, with decreased apoptosis compared to cells expressing wild-type RUNX1. Expression of Group I variant led to phenotypes similar to that of empty vector, suggesting a complete loss of RUNX1 function. In parallel, we engineered isogenic T-ALL single clones with epitope-tagged RUNX1 variant introduced at the endogenous locus via CRISPR-Cas9 mediated homology recombination. Chromatin immunoprecipitation (ChIP)-seq profiling of these cells suggested a varying degree of changes in RUNX1 binding sites across the genome as a result of the RUNX1 genetic variation. On the other hand, RNA-seq profiling identified down-regulation of genes that were activated by wild-type RUNX1, again confirming the loss-of-function effects of these variants.
Finally, we performed whole-genome seq of matched leukemia and germline samples and RNA-seq of leukemia cells in 7 T-ALL cases with RUNX1 predisposition variants. In this analysis, we observed a significant enrichment of JAK3 mutations (5 of 7 cases, 71.4%) compared to a cohort of 264 T-ALL with wild-type RUNX1 in the germline (P=3.39×10-7). By comparison, only 27.3% (3 of 7) of T-ALL with a somatic mutation in RUNX1 had concurrent JAK3 mutations in this cohort. Unsupervised clustering based on RNA-seq derived gene expression profile showed that RUNX1-mutated cases, either germline or somatic, clustered tightly with early T precursor (ETP) and near-ETP immunophenotypes.
In conclusion, we comprehensively characterized 13 RUNX1 germline variants in T-ALL, ~40% of which are frameshift or nonsense. These variants result in a loss of function, by disrupting DNA binding or deleting the transcriptional activation domain, and in some cases in a dominant-negative fashion. RUNX1 genetic variation also results in significant defects in hematopoietic cell differentiation and functions in vitro, but additional somatic lesions are most likely required for overt leukemogenesis.
Gastier Foster:Incyte Corporation: Other: Commercial Research; Bristol Myers Squibb (BMS): Other: Commercial Research. Raetz:Pfizer: Research Funding. Zweidler-McKay:ImmunoGen: Employment. Mullighan:Illumina: Honoraria, Membership on an entity's Board of Directors or advisory committees, Other: sponsored travel; Pfizer: Honoraria, Other: speaker, sponsored travel, Research Funding; AbbVie: Research Funding; Loxo Oncology: Research Funding; Amgen: Honoraria, Other: speaker, sponsored travel. Hunger:Amgen: Consultancy, Equity Ownership; Bristol Myers Squibb: Consultancy; Novartis: Consultancy; Jazz: Honoraria. Relling:Servier Pharmaceuticals: Research Funding. Loh:Medisix Therapeutics, Inc.: Membership on an entity's Board of Directors or advisory committees.

This feature is available to Subscribers Only
Sign In or Create an Account Close Modal