

In this issue of Blood, 1 study megakaryocyte (MK) heterogeneity according to ploidy levels using single cell RNA sequencing (scRNASeq) and found 3 mature MK populations with different functions: 1 regulating hematopoietic stem cells (HSCs), a second involved in proplatelet formation, and a third functioning similar to a monocyte, with innate and adaptive immune responses.
Until recently, it was believed that megakaryopoiesis was solely a cellular process that leads to platelet production from HSCs. It was assumed that this process took place inside the bone marrow, although mature MKs may transmigrate into the blood circulation. Recently, 2 new concepts have emerged: (1) MK precursors can be directly produced without mitosis from a MK-biased HSCs in stress conditions,2 and (2) megakaryopoiesis may take place in the lung parenchyma.3 However, these 2 concepts did not question that megakaryopoiesis is a linear process leading to platelet production. This year, the application of single-cell transcriptomics to MKs has completely changed this traditional view. First, it has been shown that lung MKs are mainly diploid immune cells, intermediary between dendritic cells and monocytes,4,5 and second, that during development, more particularly in the fetal liver, 3 subsets of MKs are present.6 Sun et al largely extend this last concept by confirming that MKs are a heterogeneous cell population not only in fetal liver but also in murine and human bone marrow with 4 clusters6: the first is composed of MKs undergoing cell cycle (by mitosis or endomitosis) and the 3 others correspond to different subsets of mature MKs: immune-, niche-supporting, and platelet-generating MKs. Only the platelet-generating and niche-supporting MKs reach high ploidy and large size. In contrast, most immune MKs are diploid and are the size of a monocyte. Interestingly, whereas the platelet-generating MKs are transcriptionally close both between species and during development, the niche-supporting MKs differ in the expression of genes encoding secretory and cell–cell signaling molecules, reflecting differences in HSC regulation.
The most intriguing subset is the immune MKs that have been previously identified in the lung.4 It was assumed that this immune phenotype was related to the lung environment because bone marrow MKs injected in the lung could acquire immune markers such as class II antigens.4 Here, the authors studied the functions of these marrow-derived immune MKs using the CD53 marker for isolation and showed that they behave similar to antigen presenting cells with phagocytosis properties but with some important differences between the adult and fetus. This study clearly suggests that immune MKs are indeed a specific subset of cells. Furthermore, although immune MKs may respond to thrombopoietin (TPO), they are unable to produce platelets after TPO stimulation. Notably, if immune MKs are unable to produce platelets, this subset cannot provide the explanation for why platelets have immune functions or express toll-like receptors (TLR). The likely stimulus of immune MK seems to be inflammation, because they quickly increase their number and function after lipopolysaccharide (LPS) stimulation. It would be of great interest to determine whether a link exists between immune MKs and certain dendritic cells and SLAMF7 monocytes, the precursors of fibrocytes that express myeloproliferative leukemia (MPL) protein and respond to TPO.7 Here, the authors suggest that they are different, as evidenced by markers such as CD11b. However, at the transcriptional level, if immune MKs aggregate in a different cluster than monocytes and bone marrow dendritic cells, then their transcriptional program is regulated by PU.1 and IRF8 and not by GATA1. Thus, the transcriptional program of immune MKs is closer to the mononuclear phagocyte system (MPS) than to traditional MKs.8
This innovative study raises important questions that will open new research avenues. Is megakaryopoiesis not only dedicated to the platelet production with MKs but actually much more complex with 3 different faces with distinct functions? In this study, the algorithms used for analysis of the different subsets indicate that the polyploidization MK (CD41+ cell) will first give a first branch (immune MKs) and then the common pathway will separate into 2 independent branches: the niche-supporting MKs and the platelet-generating MKs (see figure, panel A). This result would mean that only about half of mature MKs are involved in platelet production. However, in this manuscript, the authors could show that niche-supporting MKs are located close to HSCs in the marrow, whereas platelet-generating MKs are in contact with the endothelium. Furthermore, in the presence of TPO, some of the niche-supporting MKs were able to give rise to platelets. Thus, knowing that during differentiation MK may migrate in the bone marrow toward the endothelium, we cannot exclude that niche-supporting MKs are the direct precursors of platelet-generating MKs (see figure, panel B). The main differences between these 2 subsets are the cytokine transcriptional profiles, and this could be either the consequence of the MK location in the marrow or the reflection of an intrinsic program of differentiation mimicking granulopoiesis where an arrest in the synthesis of some granule proteins occurs early in differentiation. Thus, it will be important to use proteomic methodology to clearly understand what the precise relationship is between these 2 subsets. As the cycling MK cluster is also heterogeneous, it cannot be excluded that the immune MKs do not really belong to the MK lineage but to MPS. They could derive from a specific progenitor or a progenitor common with dendritic cells or with monocytes (see figure, panel C).8 Alternatively, it has been shown that inflammation induces stress megakaryopoiesis by activating stem-like megakaryocyte-committed progenitors that very efficiently produce platelets.2 Thus, it may be that this progenitor close to MK-biased HSCs can, in case of emergency, gives rise both to immune MKs, which downregulate the inflammatory process, and traditional MKs to increase platelet production (see figure, panel D).
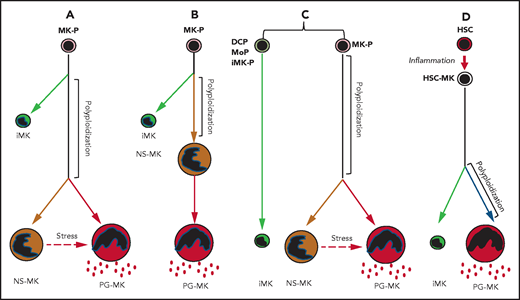
Schema of 4 algorithms giving rise to 3 different MK populations. Immune megakaryocytes (iMKs), niche-supporting megakaryocytes (NS-MKs), and platelet generating megakaryocytes (PG-MKs). The third algorithm includes the possibility of generation of iMKs from dendritic cell progenitors (DCPs), from monocytic progenitors (MoPs), or from immune MK-progenitors (iMK-Ps). The fourth algorithm summarizes the generation of iMKs and PG-MKs directly from MK-primed hematopoietic stem cell (HSC-MK) during inflammation.
Schema of 4 algorithms giving rise to 3 different MK populations. Immune megakaryocytes (iMKs), niche-supporting megakaryocytes (NS-MKs), and platelet generating megakaryocytes (PG-MKs). The third algorithm includes the possibility of generation of iMKs from dendritic cell progenitors (DCPs), from monocytic progenitors (MoPs), or from immune MK-progenitors (iMK-Ps). The fourth algorithm summarizes the generation of iMKs and PG-MKs directly from MK-primed hematopoietic stem cell (HSC-MK) during inflammation.
Finally, this paper raises an important question as to the role of TPO as an immunoregulatory cytokine. For example, thrombopoietin receptor agonists are used as treatment of immune thrombocytopenia purpura (ITP) and aplastic anemia, and their therapeutic role was thought to be caused by their role in platelet production and on HSC survival. In light of these data, could another scenario be that they also play a role in modulating the autoimmune disorder associated with ITP and some cases of aplastic anemia?
Conflict-of-interest disclosure: The authors declare no competing financial interests.

This feature is available to Subscribers Only
Sign In or Create an Account Close Modal