

TO THE EDITOR:
In rare instances, malignant lymphomas present as a body cavity–based effusion, without an identifiable tumor mass. Most frequently, this concerns primary effusion lymphoma (PEL), a human herpes virus-8 (HHV8)-positive B-cell lymphoma that typically occurs in HIV-positive patients. PEL has a characteristic phenotype, lacking pan–B-cell markers (CD19, CD20, CD79a), but is often positive for activation and plasma cell–related markers (CD30, CD38, CD138) and generally coinfected with Epstein-Barr virus (EBV). Prognosis is poor with a median overall survival of 6 months.1
Recently, HHV8-negative effusion-based lymphoma (HHV8-negative EBL) has been described that has a very different epidemiology, generally presenting in patients >70 years of age and outside the HIV context.2 The majority of these patients have an underlying condition causing fluid overload, such as chronic heart failure, renal insufficiency, or liver cirrhosis. The disease course of HHV8-negative EBL is mild, and survival is largely determined by comorbidity and high age at diagnosis. In contrast to HHV8-positive PEL, HHV8-negative EBL has a complete mature B-cell phenotype and is generally EBV-negative, similar to conventional diffuse large B-cell lymphoma (DLBCL). Therefore, the differential diagnosis with DLBCL may be challenging and largely depends on staging information, excluding the presence of disseminated disease at time of diagnosis.
Thus far, a limited number of HHV8-negative EBL cases have been reported.2-5 In these documented cases, treatment varied from drainage alone to chemotherapy and stem cell transplantation, with remarkable good outcome in those treated with drainage only. This underpins the importance of distinguishing HHV8-negative EBL from secondary effusion of DLBCL, which has a poor prognosis and requires an essentially different therapeutic approach.6 Genomic data of a limited number of HHV8-negative EBL have been reported that do not show a consistent or characteristic pattern,2-5 precluding conclusions on their role in oncogenesis. In this study, we aimed to delineate the genomic landscape of HHV8-negative EBL using established next-generation sequencing procedures.
All cytological aspiration specimens of pleural, pericardial, and abdominal effusions with a recorded diagnosis of lymphoma diagnosed between 2000 and 2017 (n = 81) were selected from the archives of the Amsterdam UMC, Vrije Universiteit Amsterdam, Department of Pathology. All secondary lymphoma effusions based on clinical staging information or histological diagnosis were excluded. Complete pathology workup was performed (supplemental Table 1, available on the Blood Web site), identifying 10 fully diagnostic cases of HHV8-negative EBL. One additional case was received from the University of Würzburg, Germany. Cell of origin was determined using the Hans and Tally algorithms.7
Genomic DNA was extracted with QIAamp DNA FFPE Tissue and Mini Kits (Qiagen, Hilden, Germany), according to the manufacturer’s recommendation. Next-generation sequencing library preparation was done using KAPA Library Preparation (KAPA Biosystems, Wilmington, MA). Single-read 50-bp shallow whole-genome sequencing for copy number analysis and paired-end 150-bp targeted sequencing with SeqCapEZ probes (Roche Nimblegen, Madison WI) for mutation and translocation analysis were performed on a HiSeq 2500 (Illumina, San Diego CA). Sequencing reads were aligned, and duplicates were removed. Copy number analysis was performed using QDNAseq,8 and LoFreq9 was used for mutation calling. For translocation detection, BreaKmer, GRIDDS, Wham, and novoBreak were used.10-13 Translocations detected by at least 2 tools were selected.
All sequence data have been deposited at the European Genome-Phenome Archive under accession number EGAS00001002743. For more detailed information, see supplemental Methods.
We studied a series of 11 cases of HHV8-negative EBL with clinicopathological features, as listed in Table 1. The mean age of patients at diagnosis was 83 years (range 60-92 years). Six patients presented with pleural effusions causing dyspnea, and 1 presented as a pericardial effusion causing a cardiac tamponade. Seven patients had a history of chronic heart failure, supporting the concept that chronic fluid overload states are an important pathogenic factor in HHV8-negative EBL. At the time of primary diagnosis, the majority of the patients were diagnosed as DLBCL. However, none of the patients were treated as such, due to comorbidity and/or personal preference because of advanced age. Six patients died of their underlying disease within 7 months; 4 patients survived for 14 to 99 months (mean 42 months), of which 3 are still alive. For 1 patient, the vital status is unknown. No patient died of lymphoma, underpinning the indolent behavior of HHV8-negative EBL. Of 10 available cases, 9 were classified as non-GCB and 1 was classified as GCB.
For 8/11 cases, genomic profiles were derived using shallow whole-genome sequencing and targeted sequencing (supplemental Figure 1). We observed a high number of mutations, including characteristic patterns of somatic hypermutation (PIM1, BCL2, KLHL14)14 in 4/8 cases, indicating a (post-) germinal center character (Figure 1). The most recurrent mutations were found in HIST1H1E and MYD88. Of the 4 MYD88 mutations, 3 affected MYD88L265P, and comutation with CD79B was found in 1 case. These aberrations are typically found in activated B-cell–like (ABC) DLBCLs and enriched in extranodal (testis, central nervous system) DLBCL.15 In 6/8 cases, truncating mutations were found in BTG1 and/or BTG2. IRF4 and SYNE1 were mutated in 4/8 cases. Other recurrently mutated genes included chromatin modifying genes (eg, CREBBP, KMT2D, MEF2B), which are more frequently found in GCB-type DLBCLs.16
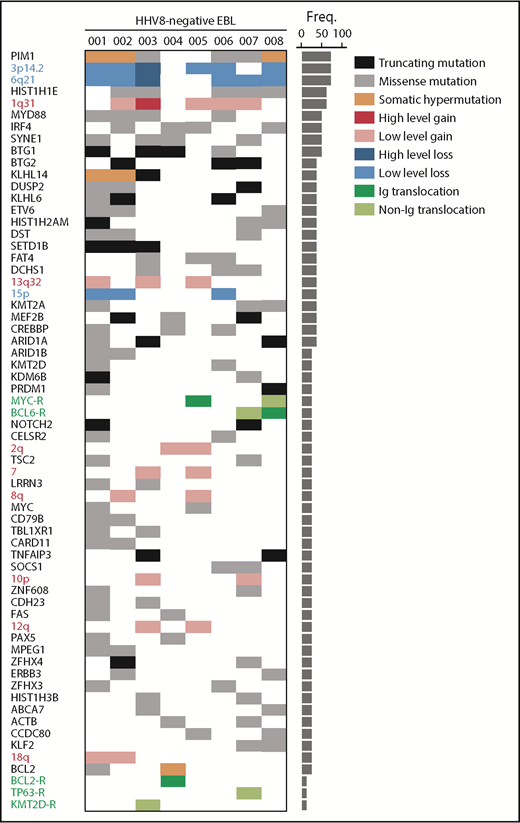
Distribution of genomic aberrations in HHV8-negative EBL. Each column represents an individual case (HHV8-negative EBL 001-008). Rows depict genomic aberrations, including mutations, CNAs, and translocations. The bar graph on the right shows the percentage of each aberration in the full cohort. Variant categories include missense mutations (gray), truncating mutations (black), low-level copy number gains (red), high-level copy number gains (dark red), low-level copy number losses (blue), high-level copy number losses (dark blue), translocations with immunoglobulin (Ig) partners (dark green), and translocations with non–immunoglobulin partners (light green). Freq., frequency.
Distribution of genomic aberrations in HHV8-negative EBL. Each column represents an individual case (HHV8-negative EBL 001-008). Rows depict genomic aberrations, including mutations, CNAs, and translocations. The bar graph on the right shows the percentage of each aberration in the full cohort. Variant categories include missense mutations (gray), truncating mutations (black), low-level copy number gains (red), high-level copy number gains (dark red), low-level copy number losses (blue), high-level copy number losses (dark blue), translocations with immunoglobulin (Ig) partners (dark green), and translocations with non–immunoglobulin partners (light green). Freq., frequency.
HHV8-negative EBL showed an overall complex copy number landscape with a mean of 33.6 copy number aberrations (CNAs) per case (range 6-133, standard deviation 34.7). This number of CNAs is similar to that reported for conventional DLBCL.17 Large CNAs that occurred in at least 2/8 cases included gains of chromosomes 1q, 2q, 7, 8q, 10p, 12q, 13q, and 18q, and losses of chromosomes 6q and 15p (Figure 1; supplemental Figure 2; supplemental Table 2). These findings are in line with classical karyotyping and comparative genomic analysis in the reported HHV8-negative EBL cases.2-5
The copy number landscape of HHV8-negative EBL showed a remarkable enrichment of focal CNAs. Specifically, focal deletions at 3p14.2 and 6q21 were present in 6/8 cases (supplemental Figure 2; supplemental Table 3). Both loci were heterozygously deleted in 5 cases and homozygously deleted in 1 case. Focal deletion of chromosome 6q21 has not been described for HHV8-positive PEL, but is observed in DLBCL, albeit at relatively lower frequency (57/304 [19%]).15 The 6q21 locus contains PRDM1/Blimp-1, a master regulator of plasma cell differentiation.18 Furthermore, PRDM1 was mutated in 2 cases that had a heterozygous deletion of 6q21, resulting in biallelic inactivation of PRDM1 in 3/8 cases (Figure 1; supplemental Figure 1). The 3p14.2 focal deletion has been described in a subset of DLBCLs (7/57 [12%]),19 but it occurred at a significantly higher frequency in HHV8-negative EBL (6/8 [75%], P = .0005). In HHV8-positive PEL, a similar frequency (85%) of FHIT deletions is reported.20 FHIT is involved in maintaining genome stability, and disruption may lead to an accumulation of genomic damage.21
The most frequent translocations were observed at the MYC, BCL2, and BCL6 loci. The presence of these translocations has been previously described in a subset of HHV8-negative EBL.2-5 We demonstrated the involvement of nonimmunoglobulin translocation partners in addition to immunoglobulin translocation partners. Furthermore, previously undescribed translocations involving TP63, EXOC2, and KMT2D were identified (supplemental Figure 1).
Analogous to findings in other clinically indolent B-cell lymphomas, we expected HHV8-negative EBL to have a relatively simple genomic landscape with low levels of mutations and CNAs.22 However, we observed that HHV8-negative EBL is characterized by a complex genomic landscape with frequent mutations, CNAs, and translocations, often found in aggressive ABC- and GCB-type DLBCL and PEL. Despite an abundance of known adverse prognostic features in HHV8-negative EBL, such as MYC translocations, the clinical course is remarkably indolent. The nonaggressive behavior of these tumor cells may be caused by the micro/macroenvironment in the primary anatomical localization, restraining progression.23 For daily practice, clinicopathologic correlation, excluding the presence of disseminated disease, remains essential.
The study and protocols to obtain human archival tissues and patient data were approved by the local ethical committee of the Amsterdam UMC/VU University Medical Center, Amsterdam (FWA00017598) and comply with the general Data Protection Act and the Code for Proper Secondary Use of Human Tissue in The Netherlands.
The online version of this article contains a data supplement.
Acknowledgments
The authors thank Paul P. Eijk (Amsterdam UMC, Vrije Universiteit Amsterdam, Department of Pathology, Amsterdam, The Netherlands) for providing administrative and material support, and Daoud Sie (Amsterdam UMC, Vrije Universiteit Amsterdam, Clinical Genetics, Amsterdam, The Netherlands) for providing genomics and computational infrastructure.
This work was supported by the Dutch Cancer Society grant KWF 2015-7925 and KWF 2012-5711.
Authorship
Contribution: M.M. designed and performed the research and analyzed and interpreted the data, and wrote the paper; M.G.M.R. analyzed and interpreted the data and wrote the paper; M.F.C.M.v.d.H. designed and performed the research and analyzed and interpreted the data; G.T.L.-d.V. performed research and analyzed and interpreted the data; R.B., P.S., and N.J.H. performed research; A.R. provided samples and reviewed the manuscript; B.Y. and D.d.J. designed the research, analyzed and interpreted the data, and wrote the paper.
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Correspondence: Daphne de Jong, Amsterdam UMC, Vrije Universiteit Amsterdam, Department of Pathology, De Boelelaan 1117, 1081 HV, Amsterdam, The Netherlands; e-mail: d.dejong2@vumc.nl.

