Key Points
HLA mismatching is associated with relapse and survival after haploidentical transplantation utilizing posttransplant cyclophosphamide.
HLA should inform the selection of haploidentical donors for transplantation.

Abstract
Hematopoietic cell transplantation from HLA-haploidentical related donors is increasingly used to treat hematologic cancers; however, characteristics of the optimal haploidentical donor have not been established. We studied the role of donor HLA mismatching in graft-versus-host disease (GVHD), disease recurrence, and survival after haploidentical donor transplantation with posttransplantation cyclophosphamide (PTCy) for 1434 acute leukemia or myelodysplastic syndrome patients reported to the Center for International Blood and Marrow Transplant Research. The impact of mismatching in the graft-versus-host vector for HLA-A, -B, -C, -DRB1, and -DQB1 alleles, the HLA-B leader, and HLA-DPB1 T-cell epitope (TCE) were studied using multivariable regression methods. Outcome was associated with HLA (mis)matches at individual loci rather than the total number of HLA mismatches. HLA-DRB1 mismatches were associated with lower risk of disease recurrence. HLA-DRB1 mismatching with HLA-DQB1 matching correlated with improved disease-free survival. HLA-B leader matching and HLA-DPB1 TCE-nonpermissive mismatching were each associated with improved overall survival. HLA-C matching lowered chronic GVHD risk, and the level of HLA-C expression correlated with transplant-related mortality. Matching status at the HLA-B leader and HLA-DRB1, -DQB1, and -DPB1 predicted disease-free survival, as did patient and donor cytomegalovirus serostatus, patient age, and comorbidity index. A web-based tool was developed to facilitate selection of the best haploidentical-related donor by calculating disease-free survival based on these characteristics. In conclusion, HLA factors influence the success of haploidentical transplantation with PTCy. HLA-DRB1 and -DPB1 mismatching and HLA-C, -B leader, and -DQB1 matching are favorable. Consideration of HLA factors may help to optimize the selection of haploidentical related donors.
Introduction
Improvements in HLA-haploidentical transplantation have increased its safety and efficacy for the treatment of life-threatening hematologic cancers.1-8 Particularly, the use of cyclophosphamide after hematopoietic cell infusion (posttransplantation cyclophosphamide [PTCy]) has substantially reduced severe acute and chronic graft-versus-host disease (GVHD) and is now the most widely used GVHD prevention strategy in haploidentical transplantation.9
A haploidentical family member shares 1 complete HLA haplotype with the patient and differs for a variable number of alleles on the nonshared haplotype. The probability that a patient has at least 1 haploidentical family member exceeds 90% because biologic parents and children by definition are haploidentical, and siblings and more distant relatives may be haploidentical.6 Haploidentical transplantation provides an expeditious therapy for patients, such as non-Caucasians, with a lower likelihood of identifying a suitable unrelated donor10-12 and patients who require urgent transplantation for high-risk disease. In 2019, the number of haploidentical transplantations performed in the US approached that of genotypically matched siblings.13
Most of the information on the importance of HLA in hematopoietic cell transplantation comes from analyses of the results of unrelated donor transplantations without PTCy. In that context, increasing numbers of donor HLA mismatches are detrimental to survival, and the clinical implications of specific HLA mismatches depend on the locus and sequence features of the mismatched alleles.14-16 The immunogenicity of a given HLA mismatch may relate to specific amino acid epitopes recognized by T cells and allogeneic HLA peptides presented by HLA.17,18 In the case of HLA-DPB1 (T-cell epitopes [TCE]), matching or permissive mismatching is associated with less acute GVHD and better survival but also higher disease recurrence (“relapse”) after unrelated transplantation.18-20 The importance of functional variation outside of the peptide-binding region is illustrated by a dimorphism unique to HLA-B leader peptides that differentially affects mortality after unrelated transplantation and cord blood transplantation and defines HLA-B mismatches that are better tolerated than others.15,16,21
In contrast, haploidentical donors are currently selected on the basis of non-HLA factors, but information on the role of HLA in haploidentical transplantation is emerging.22-26 Although the number of HLA mismatches on the nonshared haplotype has not correlated with outcomes,27 the association of lower relapse with HLA-DRB1–mismatched haploidentical donors offers new insight into the mechanisms of graft-versus-host (GVH) allorecognition.7,22,24 Whether HLA mismatches in haploidentical transplantation confer similar effects on GVHD, relapse, and mortality as those observed in unrelated donor transplantation is unknown.
We performed a retrospective analysis on the largest clinical cohort to date to study the effect of individual and combined HLA locus mismatching after haploidentical transplantation using PTCy. Our goal was to identify functional HLA characteristics that could help guide donor selection to improve the overall success of haploidentical transplantation.
Methods
Patients and HLA typing
HLA typing and clinical data were available from the Center for International Blood and Marrow Transplant Research for 1434 patients who received a haploidentical-related donor transplant with PTCy between January 2008 and December 2017 for the treatment of acute myeloid leukemia, acute lymphoblastic leukemia, or myelodysplastic syndrome (Table 1). Patients included in any prior analysis of HLA (mis)matching and outcomes were excluded from the current study. HLA-A, -B, -C, -DRB1, -DQB1, and -DPB1 alleles were typed as described and a mismatch was defined at the highest level of resolution of the allele.28 The presence of a GVH and host-versus-graft (HVG) vector of incompatibility was determined for each patient/donor HLA mismatch. Transplant pairs were defined as HLA-A, -B, -C, -DRB1, or -DQB1 mismatched in the GVH direction (bi- or unidirectional) vs matched (HLA matched or HVG unidirectional mismatch) for each locus. HLA-B mismatches were additionally defined as leader matched or leader mismatched.15 HLA-DPB1 typing was available for a total of 677 patient/donor pairs, of which 435 (64%) were retrospectively typed on available research samples from the National Marrow Donor Program repository as previously described.29 Several methods are available to define permissive and nonpermissive HLA-DPB1 mismatches.17-20 The TCE model was previously validated in unrelated donor transplants and was used to determine TCE nonpermissive or permissive mismatches and complete matches in the current study.19,20 The current study did not use HLA-DP expression to define permissive or nonpermissive mismatching.30
Biostatistical methods
The primary clinical endpoints were survival without disease recurrence (“disease-free survival”) and overall survival. Secondary endpoints were acute (grades 2-4 and 3-4) and chronic GVHD (present/absent), relapse, and mortality from causes other than relapse (“transplant-related mortality”). Cox proportional hazards models were used to compare the hazards of failure between appropriate groups. Patients were censored at date of last follow-up or second transplant. All clinical variables known to potentially affect the endpoint of interest were tested first for the affirmation of the proportional hazard assumption including donor age, patient age, race/ethnicity, disease and stage, cytomegalovirus (CMV) serostatus, time from diagnosis to transplant, transplant comorbidity index,31 donor relationship, conditioning intensity, graft type, Karnofsky performance score, and sex matching (Table 1). Factors violating the proportional hazards assumption were adjusted through stratification. Following this, stepwise forward and backward elimination procedures were performed to determine the adjusted clinical variables (with a threshold of P = .05 for both entry and retention in the model). The total number of HLA-A, -B, -C, -DRB1, and -DQB1 allele mismatches and HLA-DPB1 TCE (non)permissive mismatches, all in the GVH vector, were also examined. Patients with missing data for a particular outcome were excluded from the appropriate regression analysis. Two-sided P values were obtained from the Wald test for Cox regression models. To adjust for multiple testing of HLA variables, an overall value of P of < .01 was considered significant.
Informed consent
Informed consent was obtained from patients and donors in accordance with the Declaration of Helsinki. Protocols were approved by the institutional review boards of the National Institutes of Health Office for Human Research Protections and the National Marrow Donor Program Institutional review board. The funding agencies had no role in study design, data collection and analysis, the decision to submit the manuscript for publication, or the preparation of the manuscript.
Results
For the entire population of patients transplanted at 111 centers in this study, the 100-day cumulative incidence of grades 2-4 acute GVHD was 34.7% and 3-4 was 9.3%; the 1-year cumulative incidence of chronic GVHD was 25.8%; the 3-year cumulative incidences of nonrelapse mortality and of relapse were 19.7% and 39.1%, respectively, and the 3-year actuarial overall and disease-free survivals were 52.5% and 41.3%, respectively.
Effect of the number of mismatches on clinical outcome
Haploidentical patients and donors are matched for the 5 HLA-A, -B, -C, -DRB1, and -DQB1 alleles of the shared haplotype and may be fortuitously matched for any number of alleles on the nonshared haplotype. The majority of pairs were matched for 5/10 (928, 65%) or 6/10 (314, 22%) alleles at HLA-A, -B, -C, -DRB1, and -DQB1.
We tested the hypothesis that the number of matched alleles informs clinical outcome. Compared with patients matched for 5 alleles, the hazard ratios (HRs) of relapse were 1.06 (95% confidence interval [CI], 0.87-1.28) for HLA-6/10 matches and 1.35 (95% CI, 1.08-1.69) for HLA-7/10 and higher matches (P = .03) (supplemental Table 1). The total number of matched alleles was not significantly associated with the risks of GVHD, disease-free survival, transplant-related mortality, or overall mortality.
Locus-specific risks
To test the hypothesis that clinical outcomes differ based on the specific location (locus) of the HLA mismatch, we assessed the risks associated with patient/donor mismatching relative to matching at HLA-A, -B, -C, -DRB1, and -DQB1 individually (Figure 1; supplemental Table 2) and in multivariable analysis accounting as well for non-HLA factors (Table 2). The frequencies of HLA characteristics were similar across races/ethnicities and therefore were examined for the entire study population (Table 1).
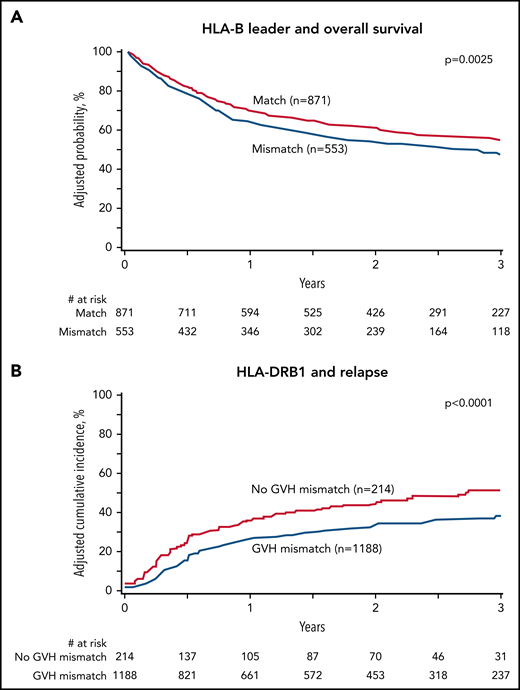

HLA-B, -DRB1, -DPB1, and -C mismatching and clinical outcome. Adjusted probabilities are derived from multivariable models. (A) HLA-B leader and overall survival. (B) HLA-DRB1 and relapse. (C) HLA-DPB1 and overall survival. (D) HLA-C and chronic GVHD. Survival models (A, C) were adjusted for comorbidities, disease type and status, recipient and donor age, recipient-donor cytomegalovirus match, time from diagnosis to transplant and stratified by graft type. Model A was also adjusted for HLA-DPB1. Model C was also adjusted for HLA-B leader. Relapse model (B) adjusted for conditioning intensity and time from diagnosis to transplant and stratified for disease type and status. Chronic GVHD model (D) was adjusted for disease type, recipient age, recipient-donor sex match, time from diagnosis to transplant, and graft type.
HLA-B, -DRB1, -DPB1, and -C mismatching and clinical outcome. Adjusted probabilities are derived from multivariable models. (A) HLA-B leader and overall survival. (B) HLA-DRB1 and relapse. (C) HLA-DPB1 and overall survival. (D) HLA-C and chronic GVHD. Survival models (A, C) were adjusted for comorbidities, disease type and status, recipient and donor age, recipient-donor cytomegalovirus match, time from diagnosis to transplant and stratified by graft type. Model A was also adjusted for HLA-DPB1. Model C was also adjusted for HLA-B leader. Relapse model (B) adjusted for conditioning intensity and time from diagnosis to transplant and stratified for disease type and status. Chronic GVHD model (D) was adjusted for disease type, recipient age, recipient-donor sex match, time from diagnosis to transplant, and graft type.
HLA-B was evaluated with both the classical definition of allele (mis)matching, which captures variation within the peptide-binding region, as well as leader (mis)matching. HLA mismatching at HLA-A, -B, or -DQB1 alone was not significantly associated with any clinical endpoint; however, when HLA-B was defined by leader match status, leader mismatching was associated with significantly lower overall survival (Figure 1A) (HR, 1.23; 95% CI, 1.07-2.18; P = .004) and higher transplant-related mortality (HR, 1.43; 95% CI, 1.13-1.82; P = .003) relative to leader matching. The effect of HLA-B mismatching was furthermore dissected from the effect of leader mismatching: compared with HLA-B–matched (ie, leader-matched) transplants, a leader-matched HLA-B allele or antigen mismatch was not associated with significantly increased risk of mortality (HR 0.93; 95% CI, 0.71-1.22; P = .62) or transplant-related mortality (HR 1.26; 95% CI, 0.56-2.85; P = 0.57). These results suggest that HLA-B leader (mis)matching, more so than classic allele and antigen (mis)matching, is a clinically relevant model of immunogenicity for HLA-B.
HLA-DRB1 mismatching in the GVH direction was associated with significantly lower relapse compared with matching (HR, 0.65; 95% CI, 0.53-0.80; P < .0001) (Table 2; Figure 1B). HLA-DPB1 TCE-nonpermissive mismatches were associated with improved overall survival relative to matches or TCE-permissive mismatches (HR, 0.59; 95% CI, 0.43-0.82; P = .002) (Table 2; Figure 1C). The risk of acute GVHD did not correlate with mismatching at any locus. The risk of chronic GVHD was higher with HLA-C mismatching compared with HLA-C matching (HR, 1.46; 95% CI, 1.17-1.83, P = .0008) (Figure 1D). The level of HLA-C expression correlated with transplant-related mortality and models adjusted for the expression level of the mismatch as previously described.32
In summary, the effects of HLA mismatching in the GVH direction depend on the mismatched locus. HLA-DRB1 mismatches decrease relapse and nonpermissive HLA-DPB1 mismatches decrease mortality. HLA-B leader matching lowers transplant-related mortality and improves overall survival. These observations suggest different underlying mechanisms through which HLA-DRB1, -DPB1, and -B mismatches influence clinical outcome after haploidentical transplantation with PTCy.
Two-locus mismatch combinations
The effects of individual mismatches on specific clinical endpoints motivated us to examine 2-locus combinations for disease-free survival and relapse (HLA-B leader with HLA-DRB1) and survival (HLA-B leader with HLA-DPB1 TCE). The known strong positive linkage disequilibrium between HLA-DRB1 and HLA-DQB1 prompted additional dissection of locus-associated effects. The limited numbers of transplants with HLA-DPB1 typing precluded meaningful analysis of 3-way effects of HLA-B with HLA-DRB1 and HLA-DPB1.
HLA-B leader with HLA-DRB1
We defined 4 transplant groups by the presence of concurrent (mis)matching for the HLA-B leader and HLA-DRB1: 152 (10.8%) leader-matched and HLA-DRB1–matched; 704 (50.2%) leader-matched but HLA-DRB1–mismatched; 62 (4.4%) leader-mismatched but HLA-DRB1–matched; and 484 (34.5%) leader-mismatched and HLA-DRB1–mismatched. The beneficial individual effects of HLA-B leader matching and HLA-DRB1 mismatching were additive for leader-matched/HLA-DRB1–mismatched patients who comprised more than half of the pairs in this unselected cohort and had superior disease-free survival and lower relapse compared with leader-mismatched/HLA-DRB1–matched patients (Figure 2). These data suggest that optimization of the HLA-B leader and HLA-DRB1 match status between patients and haploidentical donors may help to improve transplant outcomes even more for future patients.
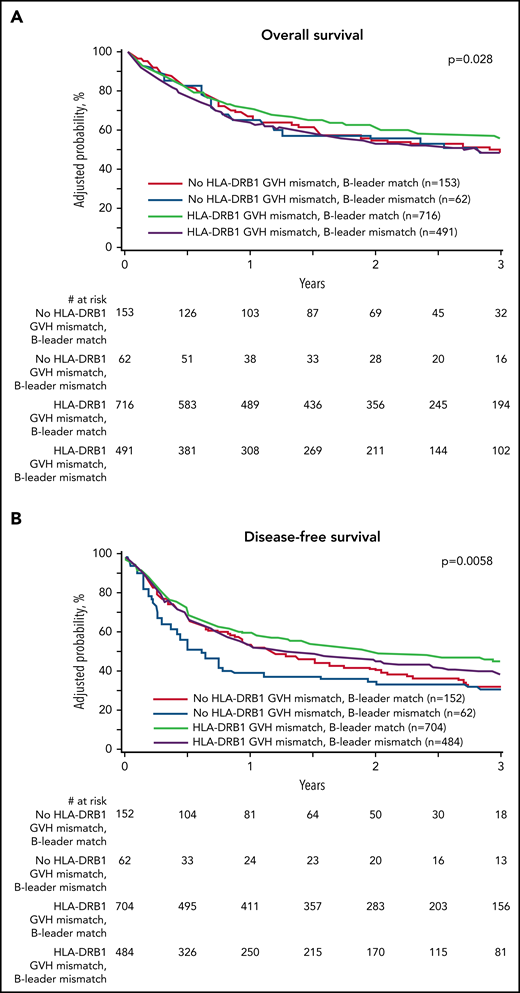
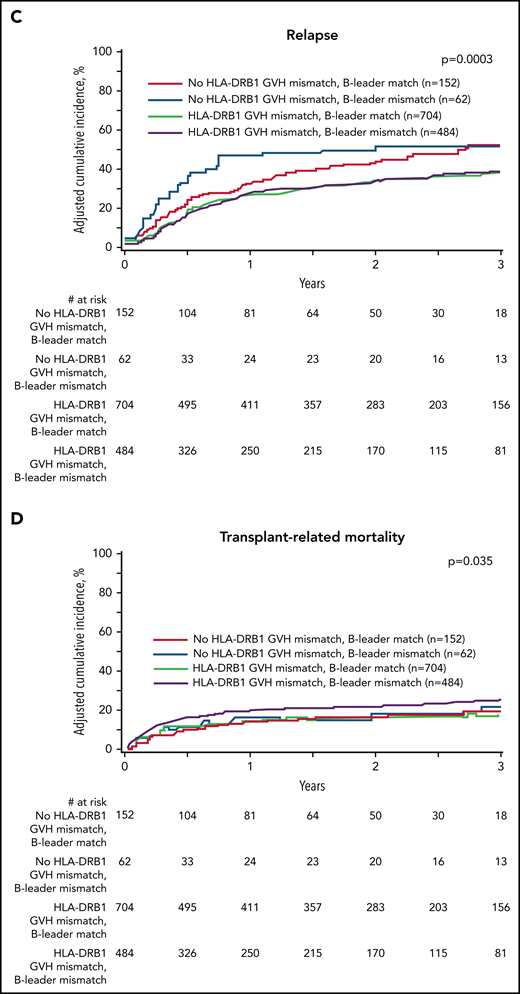
Effect of concurrent (mis)matching for the HLA-B leader and HLA-DRB1. (A) Overall survival. (B) Disease-free survival. (C) Relapse. (D) Transplant-related mortality. Probabilities are derived from multivariable models that adjusted for HLA-DPB1 mismatching. Survival model (A) was adjusted for comorbidities, disease type and status, recipient and donor age, recipient-donor cytomegalovirus match, time from diagnosis to transplant, HLA-DPB1, and stratified by graft type. Disease-free survival model (B) was adjusted for comorbidities, recipient age, recipient-donor cytomegalovirus match, donor relationship, and HLA-DPB1 and stratified by disease type and status. Relapse model (C) was adjusted for conditioning intensity and time from diagnosis to transplant and stratified by disease type and status. Transplant-related mortality model (D) was adjusted for comorbidities, conditioning intensity, disease type and status, recipient age, recipient-donor cytomegalovirus match, and recipient mean HLA-C surface expression.
Effect of concurrent (mis)matching for the HLA-B leader and HLA-DRB1. (A) Overall survival. (B) Disease-free survival. (C) Relapse. (D) Transplant-related mortality. Probabilities are derived from multivariable models that adjusted for HLA-DPB1 mismatching. Survival model (A) was adjusted for comorbidities, disease type and status, recipient and donor age, recipient-donor cytomegalovirus match, time from diagnosis to transplant, HLA-DPB1, and stratified by graft type. Disease-free survival model (B) was adjusted for comorbidities, recipient age, recipient-donor cytomegalovirus match, donor relationship, and HLA-DPB1 and stratified by disease type and status. Relapse model (C) was adjusted for conditioning intensity and time from diagnosis to transplant and stratified by disease type and status. Transplant-related mortality model (D) was adjusted for comorbidities, conditioning intensity, disease type and status, recipient age, recipient-donor cytomegalovirus match, and recipient mean HLA-C surface expression.
HLA-DRB1 with HLA-DQB1
The linkage disequilibrium between HLA-DRB1 and -DQB1 favors mismatching of both loci of the nonshared haplotype and less frequent mismatching at only HLA-DRB1 or -DQB1. Consistent with this genetic relationship, 983 (68.7%) of the transplant pairs were mismatched at both loci, 168 (11.7%) were matched at both, 49 (3.4%) were HLA-DRB1–matched/DQB1–mismatched, and 231 (16.2%) were HLA-DRB1–mismatched/DQB1–matched. HLA-DRB1–mismatched/DQB1–matched patients had the highest disease-free survival (Table 3).
HLA-B leader with HLA-DPB1
The combined effects of HLA-B leader and HLA-DPB1 (mis)matching were studied in 4 patient groups: leader-matched/DPB1 nonpermissive mismatch; leader-matched/no DPB1 nonpermissive mismatch; leader-mismatched/DPB1 nonpermissive mismatch; leader-mismatched/no DPB1 nonpermissive mismatch (supplemental Table 3). Survival was superior for leader-matched/nonpermissive mismatched compared with any other combination.
The optimal haploidentical donor and a tool for prospective donor selection
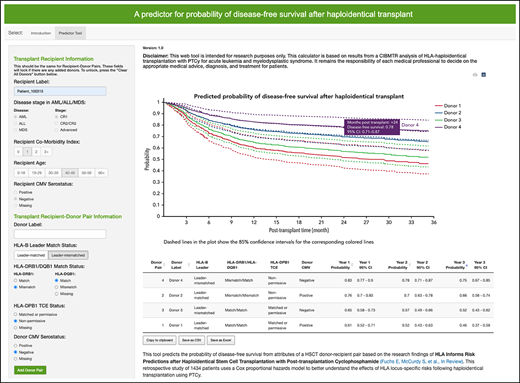
Contrasting in part to the unrelated donor setting, disease-free survival is optimized when a haploidentical donor is HLA-B leader-matched, HLA-DRB1–mismatched, HLA-DQB1–matched, and HLA-DPB1 TCE-non permissive–mismatched (Table 3). Furthermore, a CMV− donor is preferred for CMV− patients. Donor age was not associated with disease-free survival (P = .23), nor was donor relationship (P = .88). Because it is difficult to simultaneously consider all HLA and clinical factors, we developed an online calculator in R Shiny using the R-based statistical models for research purposes to aid prospective selection of haploidentical donors for patients undergoing PTCy-based haploidentical transplantation for acute leukemia or myelodysplastic syndrome (http://haplodonorselector.b12x.org/v1.0/). Based on the patient and donor information entered, 1- and 2-year disease-free survivals are estimated from the haploidentical transplants in the current study, aiding in the prioritization of candidate donors (supplemental Figure 1). The online calculator automates HLA-B leader and HLA-DPB1 TCE match assignment. Results of donor rankings can be exported for user convenience.
Discussion
The development of GVHD prophylaxis regimens that allow safe crossing of the HLA barrier has provided a life-saving therapy for patients who lack matched sibling and unrelated donors, especially patients with less common alleles and haplotypes and patients from races and ethnicities that are under-represented in unrelated donor registries. In particular, the low toxicity and ease of giving PTCy has advanced haploidentical relatives as the preferred alternative donors for many clinical centers worldwide.1,3,9 Patients often have more than 1 available haploidentical donor, and selection is currently based on non-HLA characteristics. Data on the role of HLA and transplant success are emerging and may help further refine haploidentical donor selection algorithms. Although high-resolution HLA matching clearly improves the outcome of unrelated donor transplantation without PTCy,15 the current study sought to understand the role of high-resolution HLA mismatching in the setting of haploidentical transplantation with PTCy. We found that the match status at HLA-C, -B, -DRB1, -DQB1, and -DPB1 each impact outcome and that the effects of particular combinations of loci are independent and important. Our findings have implications not only for clinical practice and donor selection but also provide new information on the potential biological mechanisms through which HLA gene products participate in the transplantation barrier.
We identified novel associations of HLA-C and -B with clinical outcome after haploidentical transplantation with PTCy and furthermore confirmed previous observations for HLA-DRB1 and -DPB1.5,27 The association of HLA-C mismatching with chronic GVHD risk mirrors its effects after unrelated donor transplantation performed with traditional methotrexate-based GVHD prevention.16 The association of HLA-C expression level with transplant-related mortality in the current haploidentical-related donor cohort mirrors the findings previously observed in unrelated donor transplantation and suggests that the level of expression of the mismatched HLA-C allele is functional.32 The lack of association of HLA mismatching with acute GVHD is striking. Potential mechanisms by which PTCy reduces acute GVHD include destruction of proliferating alloreactive T cells33 and/or preservation of regulatory T cells34 and suggests the efficacy of PTCy in ameliorating early alloimmune responses. The reason why PTCy does not abrogate the association of HLA-C mismatching with chronic GVHD is not clear but may relate to the late breakdown of thymic clonal deletion of GVH-reactive cells and their reemergence in the periphery late after PTCy.35 More detailed mechanistic studies will be required to correlate PTCy-induced molecular and cellular events with clinical outcomes.
The recent discovery of the HLA-B leader as a transplantation determinant in unrelated donor and cord blood transplantation has provided insight into the potential mechanisms underlying alloresponses.15,16,21 The current study shows that HLA-B influences clinical outcome after haploidentical transplantation through its leader and not through classical allele matching.7 In the setting of PTCy, mismatching within the peptide-binding region of HLA-B is not associated with GVHD risk, suggesting a potential contribution of pathways that involve the recognition of nonclassical class 1 ligands by natural killer (NK) cells.36,37 Although this study did not characterize the potential for NK cell alloreactivity to mediate antileukemia effects, the lack of association of HLA-C mismatch with relapse is consistent with the finding that alloreactive single–killer immunoglobulin-like receptor–positive NK cells are eliminated by PTCy and do not contribute significantly to relapse prevention.38
We confirmed the beneficial effect of HLA-DRB1 mismatching on lower relapse in an independent patient population.7,24,27 Furthermore, we found that the protective effect of HLA-DRB1 mismatching is greatest among HLA-DQB1–matched patients, suggesting that one should first consider HLA-DRB1–mismatched potential donors, then select a HLA-DQB1–matched donor if available. The dissociation of GVHD from relapse with HLA-DRB1 mismatching provides a model for understanding the mechanisms underlying graft-versus-leukemia in the setting of PTCy. We previously observed improved event-free survival among patients transplanted from HLA-DRB1 mismatched donors27 and hypothesized that donor alloreactive CD4+ T cells help effector cells to mount cytotoxic elimination of residual host leukemia. Alloreactive donor CD4+ T cells can provide help to reverse exhaustion in the patient’s tumor-specific CD8+ T cells,39-42 potentially explaining the observed dissociation of graft-versus-leukemia effects from GVHD. The differential effects of HLA-DRB1 vs HLA-DQB1 mismatching on relapse may relate to the higher cell surface expression of HLA-DRB1;43 with fewer HLA-DQB1 molecules on the cell surface, CD4+ T cells alloreactive against HLA-DQ may be unable to deliver effective help to cytotoxic effector cells. The demonstration that downregulation of HLA class II antigens from host cells is an immune escape mechanism further showcases the vigorous role of class II recognition in antileukemic recognition.24,44-46 The association of HLA-DPB1 nonpermissive T-cell epitopes with improved survival confirms previous studies and suggests mechanisms involving direct recognition of these highly immunogenic motifs of HLA-DP molecules.18-20
The selection of haploidentical donors based on HLA requires no additional testing beyond the current standards which include high-resolution definition of HLA-A, -B, -C, -DRB1, and -DQB1. Typing of HLA-DPB1 is currently included with next-generation HLA typing platforms. The assignment of the HLA-B leader furthermore requires no additional testing of the locus, and new tools are available to automate the assignment of the leader to each allele.47 When multiple haploidentical family members are available, our results may be applied to prioritize the donor providing the highest likelihood of a successful transplant. With prospective consideration of HLA-B leader matching and HLA-DRB1 mismatching, the proportion of transplants with both favorable features is likely to greatly increase the 50% observed in the current study.
Our current tool provides estimates of disease-free survival based on HLA and non-HLA characteristics in the patient and donor and will facilitate a rank ordering of preferred donors based on the best available data. The tool automates assignment of HLA-B leader genotype and HLA-DPB1 TCE status to further simplify data entry. Results of HLA antibody testing were not available to us and so were not incorporated into the selection model. In light of the association of donor-specific HLA antibodies with graft rejection and poor transplantation outcomes, a donor should not be selected if the patient has high-level antibody against that donor’s HLA molecules. Likewise, data on killer immunoglobulin-like receptor gene polymorphisms in the donor were not available to us, but their impact on donor selection should be investigated in the future as these polymorphisms may affect NK cell alloreactivity and the outcome of haploidentical transplantation.
The current study examined the effect of HLA features in patients who received a haploidentical transplant with PTCy for the treatment of acute leukemia or myelodysplastic syndrome. The study population was the largest available cohort not previously examined for HLA factors; however, the numbers of transplants matched for the HLA-B leader, DRB1, and DQB1 were limited, and our findings warrant confirmation in independent populations. Although the associations of HLA-DPB1 to outcome mirrored those observed in other cohorts, the low number of HLA-DPB1–typed pairs in the current study limited the ability to investigate synergistic effects of multilocus mismatching. Furthermore, although the current study used the TCE model to evaluate the effects of HLA-DPB1 mismatching, the role of HLA-DPB1 expression levels of outcomes of haploidentical transplantation is an area of interest for future study.30 Future analysis of patients receiving haploidentical transplantation for other blood disorders or regimens will also provide new information on the immunogenicity of HLA factors.
Given that PTCy modifies the effect of HLA mismatching on outcomes in haploidentical transplantation, further exploration of the effects of mismatching in unrelated donor transplantation utilizing PTCy is also warranted. We hypothesize that the effects of HLA matching in unrelated donor transplantation using traditional immunosuppression may shift when PTCy is employed, and future studies of the significance of HLA factors in allogeneic transplantation using PTCy are needed to reassess the “ideal” donor. This is especially relevant because it may be easier to identify optimal combinations of (mis)matches among large registries of unrelated donors than among a more restricted number of haploidentical-related donors.
In summary, the outcome of haploidentical transplantation using PTCy is improved when the donor is HLA-B leader-matched, HLA-DRB1 mismatched, HLA-DQB1 matched, and nonpermissive TCE HLA-DPB1 mismatched. These novel data provide a means to optimize donor selection and clinical outcomes for future patients.
Acknowledgments
This work was supported by grants AI069197 (E.W.P., Y.-T.B., T.W., and C.M.), CA100019 (E.W.P.), and CA218285 (E.W.P.) from the National Institutes of Health (NIH), National Cancer Institute (NCI). The Center for International Blood and Marrow Transplant Research (CIBMTR) is supported primarily by Public Health Service U24CA076518 from the NIH/NCI, the NIH/National Heart, Lung and Blood Institute (NHLBI), and the NIH/National Institute of Allergy and Infectious Diseases (NIAID), HHSH250201700006C from the Health Resources and Services Administration (HRSA), and N00014-20-1-2705 and N00014-20-1-2832 from the Office of Naval Research. Additional federal support is provided by R01AI128775, R01HL130388, and the Biomedical Advanced Research and Development Authority (BARDA). Support is also provided by Be the Match Foundation, the Medical College of Wisconsin the National Marrow Donor Program, and from the following commercial entities: Actinium Pharmaceuticals, Inc; Adienne SA; Allovir, Inc; Amgen, Inc; Angiocrine Bioscience; Astellas Pharma US; bluebird bio, Inc; Bristol Myers Squibb Co; Celgene Corp; CSL Behring, CytoSen Therapeutics, Inc.; Daiichi Sankyo Co, Ltd; ExcellThera; Fate Therapeutics; Gamida-Cell, Ltd; Genentech, Inc; Incyte Corporation; Janssen/Johnson & Johnson; Jazz Pharmaceuticals, Inc; Kiadis Pharma; Kite, a Gilead Company; Kyowa Kirin; Legend Biotech; Magenta Therapeutics; Merck Sharp & Dohme Corp; Millennium, the Takeda Oncology Co; Miltenyi Biotec, Inc; Novartis Pharmaceuticals Corporation; Omeros Corporation; Oncoimmune, Inc; Orca Biosystems, Inc; Pfizer, Inc; Pharmacyclics, LLC; Sanofi Genzyme; Stemcyte; Takeda Pharma; Vor Biopharma; and Xenikos BV.
The views expressed in this article do not reflect the official policy or position of the National Institutes of Health, the Department of the Navy, the Department of Defense, Health Resources and Services Administration, or any other agency of the US Government.
Authorship
Contribution: E.J.F., S.R.M., and E.W.P. designed the study; E.J.F. and E.W.P. drafted the manuscript; T.W. performed statistical analysis; and all authors assembled the data, critically reviewed and edited the manuscript, and approved the final version.
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Correspondence: Effie W. Petersdorf, Fred Hutchinson Cancer Research Center, Division of Clinical Research, 1100 Fairview Avenue North, Seattle, WA 98109; e-mail: epetersd@fredhutch.org.
The final analysis dataset will be posted to the CIBMTR website at https://www.cibmtr.org/ReferenceCenter/PubList/PubDsDownload/Pages/default.aspx.
The online version of this article contains a data supplement.
There is a Blood Commentary on this article in this issue.