

Regulated proteolysis of the tolerogenic matrix proteoglycan versican (VCAN) through the actions of ADAMTS-proteases, is associated with enhanced CD8+ infiltration in both hematopoietic and solid tumors. However, it is unclear whether the enhanced CD8+ infiltration results from proteolysis-mediated depletion of precursor VCAN at the tumor site or from generation of bioactive proteolytic fragments ("matrikines") (e.g., the 441-aa N-terminal fragment of V1-VCAN isoform, versikine). We have previously shown that versikine promotes Batf3-dendritic cell (DC) generation from FLT3L-mobilized bone marrow (BM) progenitors in vitro. However, the effects of versikine in DC homeostasis in the tumor microenvironment in vivo are unknown.
To investigate the effects of versikine in DC homeostasis in vivo, we utilized the first Ras-driven myeloma (MM) model (VQ model- Rajagopalan et al., Blood 132:1006, 2018) as well as transplantable solid tumor models in both C57BL/6J (LLC lung carcinoma) and Balb/c (4T1 mammary carcinoma) backgrounds. Tumor cells were stably engineered to secrete HA-tagged versikine vs. empty-vector (EV) controls.
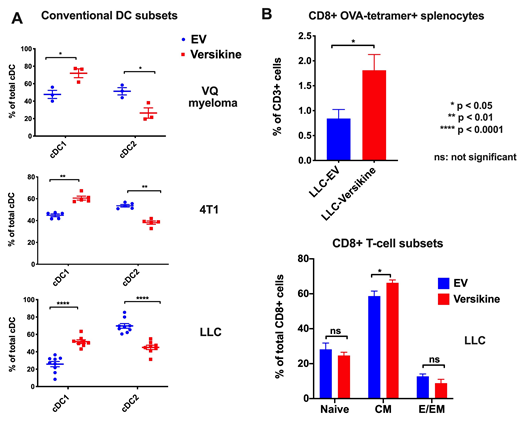
EV-VQ or versikine-VQ myeloma cells were implanted intracardiacally into C57BL/6J syngeneic recipients and mice were monitored until they developed myeloma-related end-organ damage (hindlimb paralysis). Both groups of mice were paralyzed at similar rates. Intratumoral conventional DCs (CD138-CD45+, CD11chi,MHC IIhiLy6C-, CD64-) clustered into two populations: cDC1 (Batf3-DC: CD24hi,CD11blo), a subset with crucial activity in cross-priming anti-tumor CD8+ T cells, and cDC2 (CD24lo, CD11bhi). Versikine enhanced intratumoral Batf3-DC frequency/infiltration, while cDC2 levels were diminished in versikine-VQ BM (Figure 1A)(Batf3-DC: 48% in EV-VQ vs. 72% in versikine-VQ, p-value= 0.0246; cDC2: 52% in EV-VQ vs. 28% in versikine-VQ, p=0.0312). Monocytic-derived DC (Mo-DC: CD11chi, MHC IIhi, Ly6C+, CD64+) frequency remained unchanged.
Versikine's effects were replicated in 2 solid tumor models. Versikine-expressing tumors were characterized by significantly enhanced Batf3-DC infiltration (Fig. 1A, p-value= 0.0079 for 4T1 model and <0.0001 for LLC model), whereas cDC2 numbers were diminished (p-value: 0.0079 and <0.0001 respectively). Adoptive transfer of CD45.2+ pre-DC (SIRPaint, FLIT3+, CD11c+, MHC II-, Celltrace+) in LLC-EV and LLC-versikine tumors in CD45.1+ recipients did not show any differences in 3-day differentiation potential of DC precursors, implicating other mechanisms to explain the steady-state imbalance in DC subset frequencies.
To examine whether versikine's effects on the intratumoral DC milieu in vivo could be therapeutically harnessed, we compared responses to STING agonist therapy between versikine-expressing and EV tumors. LLC-EV-OVA and LLC-versikine-OVA (ovalbumin, a model antigen) -expressing tumors received therapeutic intratumoral injections of DMXAA, a murine STING agonist. Analysis of splenocytes 5 days later showed a significant increase in the frequency of OVA antigen-specific, CD8+ (MHCI:SIINFEKL tetramer+) splenocytes in LLC-versikine-bearing animals (Figure 1B). Interestingly, there was a marked increase in total central memory T splenocytes (TCM) (CD62LhiCD44hi) harvested from LLC-versikine tumor-bearing mice.
We conclude that versikine influences the DC milieu in the tumor bed with promotion of intratumoral cross-presenting Batf3-DC and depletion of the cDC2 subset. Our findings highlight an unappreciated facet of immune regulation of the tumor microenvironment through matrix proteolytic fragments ("matrikines"). Whereas detection of native VCAN proteolysis on myeloma biopsies (see abstract by Dhakal et al.in this meeting) portends adverse outcomes likely due to the tolerogenic effects of accumulated precursor VCAN at the tumor site, therapeutic use of the isolated, purified fragments may promote tumor innate sensing and effector priming. VCAN-matrikines, through their effects on intratumoral Batf3-DC and antigen-specific CD8+ T cell infiltration, may potentiate in situ vaccination strategies across diverse hematopoietic and solid tumor types.
No relevant conflicts of interest to declare.

This feature is available to Subscribers Only
Sign In or Create an Account Close Modal