

Introduction
Enhancer of Zeste Homolog 2 (EZH2), the catalytic domain of Polycomb Repressive Complex 2 (PRC2), mediates the repressive mark of trimethylation of histone H3 lysine 27 (H3K27me3). The EZH2 gene is located on chromosome (chr.) 7q36.1 (frequently deleted in AML/MDS). Its role in tumorigenesis appears to be context-dependent since both EZH2 overexpression and loss of function are associated with different types of cancer. In patients (pts) with MDS or MDS/MPN, loss-of-function EZH2 mutations (mut) are recurrently found, associated with a poor prognosis, and EZH2 is considered a tumor suppressor gene (TSG). In AML, the incidence of EZH2-mut is lower and less well-studied. We wished to determine the effects of EZH2-mut and copy number (CN) on EZH2 expression, and the consequences for H3K27me3 levels in vivo.
Methods
Fifty-eight pts (51 AML, 3 MDS, 4 MDS/MPN), median age 63.5 years (yr, range 20-86, almost all diagnosed between 2015 and 2017), with available sequencing data, EZH2 CN status and EZH2 expression data were studied. Mutation status was determined by next-generation sequencing (NGS) including a panel of 54 genes (Illumina Myeloid NGS panel). Metaphase cytogenetics and/or fluorescence in situ hybridization and single nucleotide polymorphism arrays (selected pts) were conducted to determine EZH2 CN status. Protein expression of EZH2 and H3K27me3 was assessed semi-quantitatively by immunohistochemistry (IHC) on formalin-fixed EDTA-decalcified paraffin-embedded bone marrow (BM) core biopsies. The Kruskal-Wallis test and Dunn's multiple comparison test were used to address whether EZH2 protein expression was associated with EZH2-mut and CN.
Results
The EZH2 gene showed mutations (mostly missense or nonsense, median variant allele frequency (VAF) 45%, range 7-63%) in 13/58 pts and was unmutated in 45/58 pts. EZH2-mut pts had a median of 4 mutations. Additional mutations were most frequently found in ASXL1 (10/13; median VAF 35%, range 19-48%), less frequently in TET2, RUNX1, STAG2 (3/13 each), NRAS (2/13), and DNMT3A (1/13). In contrast, EZH2-wt pts had a median of 2 mutations, most frequently in DNMT3A (12/45), followed by NRAS (10/45), IDH1 (9/45), FLT3 (8/45), and NPM1 (7/45). Regarding chr. 7, 43 pts had no detectable deletion, 15 had 7q-/-7. Notably, the incidence of EZH2-mut was similar in pts with 7q-/-7 lesions (3/15, i.e. 20%) and pts with normal chr. 7 (10/43, 23%).
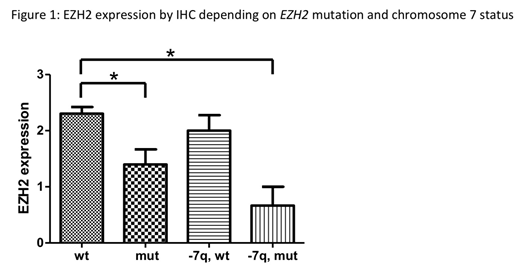
EZH2 expression in neoplastic BM cells ranked from no (score 0) to strong expression (score 3). While the hematopoietic BM cells of healthy donors usually showed a moderate EZH2 expression (score 2), in our cohort the score was 0 in 3, 1 in 13, 2 in 23, and 3 in 19 pts, respectively. We next asked whether EZH2 protein expression differed between pts depending on EZH2-mut and chr. 7 status. In Figure 1, 4 subgroups are depicted, showing highest expression in pts with EZH2-wt and either no chr. 7 abnormalities (group A, n=33) or 7q-/-7 (group C, n=12). In comparison, expression was significantly reduced in pts with EZH2-mut and no chr. 7 abnormalities (group B, n=10), and lowest in pts with EZH2-mut and chr. 7 abnormalities (group D, n=3), p<0.05 (indicated by asterisks).
Since functional EZH2 protein is necessary for the trimethylation of H3K27, the presence of this histone mark was also determined semi-quantitatively by IHC in 40 pts. H3K27me3 levels were variable, and a test for a possible association between EZH2 and H3K27me3 levels by linear regression analysis did not show an association (R²=0.13).
Sixty-two % of EZH2-mut (median age 71 yr) and 51% of EZH2-wt pts (median age 61 yr) received allografting. Median overall survival in EZH2-mut pts was 16.1 months compared to 23.5 in EZH2-wt pts (p=0.16).
Conclusions
Inactivating EZH2 mutations are infrequent events in AML. In our study, they were strongly associated with mutations of ASXL1 (also involved in PRC2 function). We did not observe overrepresentation of EZH2-mut in 7q-/-7 pts, in line with Bejar et al. (N Engl J Med 2011); this is in contrast to the frequent cooperative events between TSG mutations and deletions, e.g. of TP53. Functionally, EZH2 mutations appeared to have a stronger effect on decreased EZH2 expression than hemizygosity. Global H3K27me3 levels were not altered by EZH2 reduction, which could be due to compensatory upregulation of EZH1, supporting the clinical development of EZH1/2 inhibition.
No relevant conflicts of interest to declare.

This feature is available to Subscribers Only
Sign In or Create an Account Close Modal