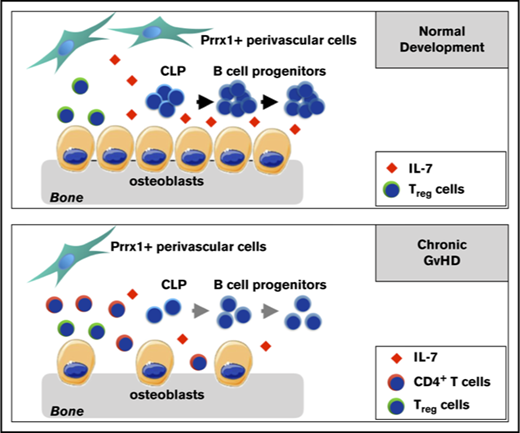
Key Points
BM B-cell development is disrupted in the BO mouse model of cGVHD due to aberrations in the B-cell progenitor niche.
Inhibition of conventional T-cell activity by donor Tregs enhanced the recovery of BM B-cell progenitors and reduced cGVHD severity.

Abstract
Chronic graft-versus-host disease (cGVHD) causes significant morbidity and mortality in patients after allogeneic bone marrow (BM) or stem cell transplantation (allo-SCT). Recent work has indicated that both T and B lymphocytes play an important role in the pathophysiology of cGVHD. Previously, our group showed a critical role for the germinal center response in the function of B cells using a bronchiolitis obliterans (BO) model of cGVHD. Here, we demonstrated for the first time that cGVHD is associated with severe defects in the generation of BM B lymphoid and uncommitted common lymphoid progenitor cells. We found an increase in the number of donor CD4+ T cells in the BM of mice with cGVHD that was negatively correlated with B-cell development and the frequency of osteoblasts and Prrx-1–expressing perivascular stromal cells, which are present in the B-cell niche. Use of anti-DR3 monoclonal antibodies to enhance the number of donor regulatory T cells (Tregs) in the donor T-cell inoculum ameliorated the pathology associated with BO in this model. This correlated with an increased number of endosteal osteoblastic cells and significantly improved the generation of B-cell precursors in the BM after allo-SCT. Our work indicates that donor Tregs play a critical role in preserving the generation of B-cell precursors in the BM after allo-SCT. Approaches to enhance the number and/or function of donor Tregs that do not enhance conventional T-cell activity may be important to decrease the incidence and severity of cGVHD in part through normal B-cell lymphopoiesis.
Introduction
Allogeneic bone marrow (BM) or stem cell transplantation (allo-SCT) is the preferred treatment of patients with relapsed/refractory or high-risk hematolymphoid malignancies otherwise not responsive to salvage treatment. The greater utilization of allo-SCT is complicated by the occurrence of graft-versus-host disease (GVHD) mediated by the recognition of major or minor major histocompatibility complex disparities in the host by donor T cells. GVHD can be classified into an inflammatory process mediated by cytolytic donor T cells and the generation of proinflammatory cytokines termed acute GVHD (aGVHD) and a profibrotic process mediated by donor T cells, macrophages, and B cells termed chronic GVHD (cGVHD).1-3
Overall survival and quality of life are decreased for patients who develop significant cGVHD.4,5 This has led to increased attention to the treatment and pathophysiology of cGVHD. Immune mechanism studies in cGVHD are limited by the paucity of animal models that mimic the clinical findings in patients with cGVHD. Our group and collaborators have used a model in which recipient mice develop B-cell–dependent lung dysfunction after conditioning therapy with total body irradiation and cyclophosphamide6 and transplantation of major histocompatibility complex–mismatched donor T cells and BM. Recipient mice developed lung dysfunction consistent with bronchiolitis obliterans (BO) as well as pathology in the liver, tongue, salivary gland, and thymus with fibrotic changes noted in these tissues; these findings are consistent with pathological changes in patients with cGVHD.7 Using this model, we have shown that antibody deposition is required for lung and liver pathology. Additionally, germinal center (GC) reactions in which T follicular helper cells interact with B cells in the GC leading to B-cell proliferation, differentiation, and antibody class switching are critical to the pathogenesis of cGVHD in this model.8
Previous work has demonstrated that somatic mutation of B cells, an important process for affinity maturation of antibody mediated by follicular helper T cells, is impaired in BM transplant recipients with cGVHD.9,10 Furthermore, cGVHD is associated with increased B-cell receptor activation and signaling in donor B cells.8,11 In addition, there was a significant negative association between the number of TdT+ B-cell precursors in the BM on day 30 after allo-SCT and the development of cGVHD in patients.12 Impaired development of donor B cells in the BM has been shown previously in different murine models of aGVHD13 and in patients with aGVHD and cGVHD.14,15 However, the mechanisms responsible for the impaired development of B cells during cGVHD have not been shown previously. Thus, we were interested in evaluating the effects of cGVHD on B-cell progenitors as they undergo differentiation from common lymphoid progenitors (CLPs) to immature B cells. This may be critical to the pathogenesis of cGVHD, as impairment of B-cell development in the BM can lead to the generation of highly autoreactive B cells in the peripheral compartment.16
To evaluate B-cell lymphopoiesis, we characterized B-cell development in the BO model of cGVHD. We demonstrated a decrease in the number of CLP, pro-, pre-, and immature B cells in the BM of mice that develop cGVHD with decreased expression of a critical lineage-specific factor in the BM. Abnormal B-cell development was mediated, in part, by donor CD4+ T cells infiltrating the BM and was improved by increasing the number of donor regulatory T cells (Tregs), which correlated with improvement in B-cell development and diminished cGVHD pathology.
Materials and methods
Mice
C57BL/6J (H2b), B10.BR (H2k), and FoxP3-GFP mice on C57BL/6J background were purchased from The Jackson Laboratory. To characterize the perivascular stromal cells in the BM, we generated Prrx1-Cre-eYFP (Prrx1+YFP+) animals using R26R-eYFP and Prrx1-Cre strains (The Jackson Laboratory). All experiments were performed in accordance with protocols approved by the University of North Carolina Institutional Animal Care and Use Committee.
Transplantation
BM transplantation was performed as described previously7 with recipient mice receiving 120 mg/kg cyclophosphamide intraperitoneally (i.p.) on days −3 and −2 followed by 7 Gy total body irradiation on day −1 (X-RAD 320, Precision X-ray). Donor-derived BM was depleted of T cells using anti-CD90.2 antibody–coated ferromagnetic beads (TCD BM), followed by the negative selection of the labeled cells using magnetic-activated cell sorting columns (Miltenyi Biotec). Splenic T cells were prepared using a total T-cell isolation column kit (Cedarlane Laboratories) or EasySep mouse T cell isolation kit (Stem Cell Technologies). On day 0, recipients were injected with 107 TCD BM with or without 5 × 104 splenic T cells. Animals were monitored 3 times a week and scored for clinical symptoms of GVHD using a semiquantitative system described previously.17
For the experiments using in vivo–expanded Tregs, donor FoxP3-GFP animals were injected i.p. with 0.5 mg/kg agonistic anti-DR3 monoclonal antibody (4C12; BioLegend) 4 days before transplantation. Splenic T cells were isolated using the method described above. Expansion of Tregs was evaluated by staining for CD4 and GFP fluorescence using flow cytometry. For the experiments involving coinfusion of purified Tregs, cells were isolated from spleen using a Treg isolation kit (Miltenyi Biotech) according to the manufacturer’s protocol. For these transplants, irradiated recipient mice received 2 × 104 Tregs given with 5 × 104 conventional T cells (Tcons) and TCD BM. Transplants evaluating for the development of aGVHD were performed with recipient mice receiving 750 cGy irradiation followed by transplantation with 5 × 105 splenic donor T cells plus TCD donor BM.
Mouse pulse oximetry
Respiratory function of mice was assessed on day 30 using a MouseOx Plus with thigh sensor (Starr Life Sciences). Oxygen saturation levels (SpO2) measurements were recorded for at least 30 seconds, and the average of all SpO2 measurements during this time was used as a data point for an individual mouse.18
Flow cytometry and cell sorting
BM cells were harvested from tibiae and femurs of mice on day 30. Single-cell suspension preparations from the bones of one hind leg were stained with fluorochrome-labeled antibodies (see supplemental Methods). Samples were analyzed using BD LSRFortessa (BD Biosciences). Cell sorting of the labeled BM pre-pro–B, pro-B, and pre-B cells was performed using MoFlo Legacy (Beckman Coulter) or BD FACSAria II (BD Biosciences).
Osteoblast isolation
Osteoblasts were isolated from the tibiae and femurs after BM removal. Bones were then mechanically broken using a mortar and pestle. Resulting fragments were thoroughly washed with phosphate-buffered saline and enzymatically digested by incubating in 0.1% collagenase and 0.05% dispase for 1 hour at 37°C.19 Undigested bone fragments were removed by filtration. Cells released by enzymatic treatment were then centrifuged and resuspended in staining buffer.
qRT-PCR
Sorted BM B-cell progenitor cells were lysed using TRIzol (ThermoFisher). Total RNA extraction was performed with the RNeasy Micro Kit (QIAGEN) according to the manufacturer’s instructions. Eluted RNA was reverse transcribed with SuperScript III and random hexamers (ThermoFisher) using the manufacturer’s protocol. Real-time quantitative PCR (qRT-PCR) was performed using TaqMan gene expression assays as described by the manufacturer (Pax5, Mm00435501_m1; Ebf1, Mm00432948_m1; Hprt, Mm03024075_m1). All experiments were performed in triplicate. Differences in the complementary DNA input were compensated by normalizing gene expression; relative expression of genes of interest was calculated using ∆∆Ct method.
Histopathology
Lung and liver tissue from perfused animals was harvested on day 30. For evaluation of the aGVHD model, small intestine and colon tissues were harvested on day 17. Samples were fixed in 10% phosphate-buffered saline–buffered formaldehyde for 48 hours. Further tissue processing, hematoxylin and eosin, and Masson’s trichrome staining was performed by the UNC Animal Histopathology and Laboratory Medicine Core facility. Samples were analyzed using Olympus BX61 microscope equipped with Hammatsu ORCA RC and QImaging RETIGA 4000R cameras controlled by Improvision’s Velocity software or Nikon Eclipse Ti microscope with DS-U3 camera using NIS Elements BR v4.50 software.
Cytokine analysis
Levels of tumor necrosis factor α (TNF-α) and interferon-γ (IFN-γ) in homogenized tissues were analyzed using an enzyme-linked immunosorbent assay (ThermoFisher) according to manufacturer’s protocol.
Statistical analyses
Survival differences were evaluated using a Mantel-Cox log-rank test. Survival curves were generated using the Kaplan-Meier method. Differences in GVHD scores and weight changes were determined using a Student t test with Holm-Sidak correction for multiple comparisons. Group comparisons of flow cytometry, pulse oxymetry and enzyme-linked immunosorbent assay data were performed using Mann-Whitney U test. P ≤ .05 was considered statistically significant.
Data-sharing statement
For original data, please contact O.V.K. (kolupaev@e-mail.unc.edu).
Results
Pathology in cGVHD target organs and lung dysfunction in B6→B10.BR Mouse Model
To demonstrate that we could recapitulate the BO model of cGVHD developed at the University of Minnesota,7,20 we characterized liver and lung pathology in recipient B10.BR mice that received sublethal irradiation and cyclophosphamide i.p. followed by TCD BM and a low dose of total splenic B6 T cells. Control animals received TCD BM without T cells. Recipient mice receiving donor T cells exhibited multiorgan pathology, with increased collagen deposition in the liver and lung (Figure 1A) compared with control animals receiving TCD BM alone. To clinically evaluate lung dysfunction in this model, we measured arterial oxygen saturation, which is diminished in animals with significant obstructive and/or restrictive lung dysfunction. Animals receiving TCD BM and donor T cells demonstrated significantly lower arterial oxygen saturation measured by pulse oximetry compared with recipient mice receiving TCD BM alone (Figure 1B). In addition to increased fibrosis of the target organs in the cGVHD group, we also detected an increase in the number of lymphocytes in the perivascular regions of the liver (Figure 1C).
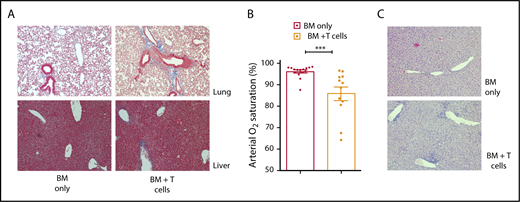
Lung and liver pathology and lung dysfunction in B6→B10.BR mouse model of cGVHD. (A) Collagen deposition (blue staining) in lung and liver of control animals (left column) and cGVHD animals (right column) (Masson’s trichrome staining, Nikon Eclipse Ti microscope, original magnification ×200). (B) Arterial oxygen saturation was measured using pulse oximetry in cGVHD animals; ***P < .001 (Mann-Whitney U test); n = 15 and n = 12 for the BM-only and BM plus T cells groups, respectively. Bars represent standard error of the mean. (C) Lymphocyte infiltration in the liver of control animals (top panel) and cGVHD animals (bottom panel) (hematoxylin and eosin staining, bright-field microscopy using Nikon Eclipse Ti microscope, original magnification ×100).
Lung and liver pathology and lung dysfunction in B6→B10.BR mouse model of cGVHD. (A) Collagen deposition (blue staining) in lung and liver of control animals (left column) and cGVHD animals (right column) (Masson’s trichrome staining, Nikon Eclipse Ti microscope, original magnification ×200). (B) Arterial oxygen saturation was measured using pulse oximetry in cGVHD animals; ***P < .001 (Mann-Whitney U test); n = 15 and n = 12 for the BM-only and BM plus T cells groups, respectively. Bars represent standard error of the mean. (C) Lymphocyte infiltration in the liver of control animals (top panel) and cGVHD animals (bottom panel) (hematoxylin and eosin staining, bright-field microscopy using Nikon Eclipse Ti microscope, original magnification ×100).
Taken together, these data indicate that we can duplicate the fibrotic manifestations consistent with cGVHD found in B10.BR recipients after the administration of TCD BM with low-dose splenic T cells from B6 donors.
Abnormalities in B-cell development in cGVHD mice
To evaluate the development of B cells in the BM after transplant, we analyzed CLP cells defined as Lin−c-KitintSca-1int IL-7R+Flt3+ cells. Animals that developed cGVHD showed a significant (twofold) reduction in the frequency and total number of CLP cells (Figure 2A-B) compared with control animals receiving TCD BM alone. We next examined whether cGVHD affected the committed B-cell lineage precursors in the BM. In addition to lower total number of CLPs, the frequency and overall number of pro-B, pre-B, and immature B cells was decreased in B10.BR mice receiving TCD B6 BM and T cells compared with the BM-only group (Figure 3A-C). Interestingly, despite this reduction, the frequency of the earliest lineage of committed B-cell progenitors in the BM, pre-pro–B cells (B220+CD43+CD19−IgM−CD11b−), was only slightly decreased in recipients of B6 T cells compared with the control group (1.35% ± 0.087% vs 1.09% ± 0.099%, P = .018); the total number of pre-pro B cells were not significantly different between the 2 groups (Figure 3A,C). The lack of a difference in overall cell number between mice receiving donor T cells and those receiving only donor BM was due to a significant increase in the frequency of pre-pro–B cells in the B220+ population (6.2% ± 0.33% vs 14.6% ± 2.75%) in mice receiving TCD BM and T cells. As previously reported for this model, we also found a decrease in the frequency of follicular B cells (IgMintAA4.1−CD23+CD21int) in the spleen of cGVHD animals20 (supplemental Figure 1). It is important to point out that the observed changes were limited to the CLP and B lymphoid lineage, as both the frequency and total number of multipotent Lin−Sca-1+c-Kit+ (LSK) cells were not affected by the infusion of B6 T cells (supplemental Figure 2). Thus, cGVHD is associated with a significant decrease in CLP cells and committed B-cell progenitors in the BM.
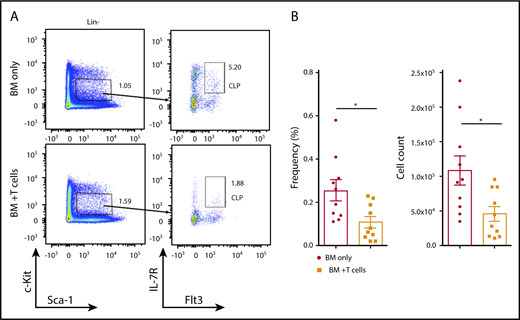
Reduction of CLP cells in the BM of cGVHD animals. B10.BR mice were transplanted with BM only or BM and T cells from B6 donor mice, and BM cells from tibiae and femurs were harvested on day 30. (A) Representative flow cytometry plots demonstrating frequency of CLP cells in the Lin−c-KitintSca-1int population in the BM of transplanted animals. (B) Frequency (left) and total number (right) of donor-derived CLP cells isolated from BM of transplanted mice. Combined data from 3 independent experiments are shown. *P < .05 (Mann-Whitney U test); n = 10 in each group. Bars represent standard error of the mean.
Reduction of CLP cells in the BM of cGVHD animals. B10.BR mice were transplanted with BM only or BM and T cells from B6 donor mice, and BM cells from tibiae and femurs were harvested on day 30. (A) Representative flow cytometry plots demonstrating frequency of CLP cells in the Lin−c-KitintSca-1int population in the BM of transplanted animals. (B) Frequency (left) and total number (right) of donor-derived CLP cells isolated from BM of transplanted mice. Combined data from 3 independent experiments are shown. *P < .05 (Mann-Whitney U test); n = 10 in each group. Bars represent standard error of the mean.
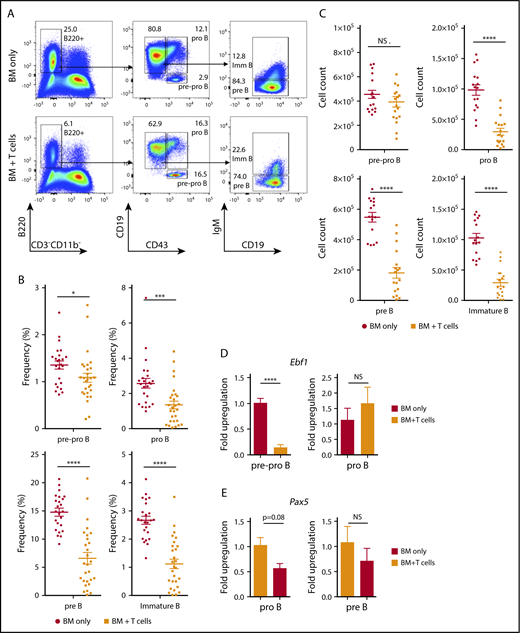
Impaired BM B-cell development in cGVHD. B10.BR mice were transplanted with BM only or BM and T cells from B6 donor mice, and BM cells from tibiae and femurs were harvested on day 30. (A) Representative flow cytometry plots demonstrating frequency of donor-derived pre-pro–B, pro-B, pre-B, and immature B cells in the BM of transplanted animals. The numbers indicate the frequency of cells found in the gated (not total) population. (B-C) Frequency of cells found from all cells evaluated in the BM (B) and total number (C) of donor-derived pre-pro–B (B220+CD43+CD19−IgM−), pro-B (B220+CD43+CD19+IgM−), pre-B (B220+CD43−CD19+IgM−), and immature B cells (B220+CD43−CD19+IgM+) isolated from the BM of transplanted mice. Combined data from 5 independent experiments are shown. *P < .05, ***P < .001, ****P < .0001 (Mann-Whitney U test); n = 24 (16 for overall number of cells) and n = 28 (18 for overall number of cells) for the BM only and BM plus T cells groups, respectively. (D) Expression of Ebf1 in sorted pre-pro–B and pro-B progenitors isolated from the BM-only control group (open bars) and cGVHD group (filled bars). Pooled data from 2 independent experiments are shown. ****P < .0001 (unpaired Student t test); n = 6 for each group; sorting purity was >95% and >90% for pre-pro–B and pro-B progenitors, respectively. (E) Expression of Pax5 gene in sorted pro-B and pre-B progenitors isolated from the BM-only control group (open bars) and cGVHD group (filled bars); n = 3 for each group, >90% and >95% for pro-B and pre-B progenitors. Bars represent standard error of the mean. NS, not significant.
Impaired BM B-cell development in cGVHD. B10.BR mice were transplanted with BM only or BM and T cells from B6 donor mice, and BM cells from tibiae and femurs were harvested on day 30. (A) Representative flow cytometry plots demonstrating frequency of donor-derived pre-pro–B, pro-B, pre-B, and immature B cells in the BM of transplanted animals. The numbers indicate the frequency of cells found in the gated (not total) population. (B-C) Frequency of cells found from all cells evaluated in the BM (B) and total number (C) of donor-derived pre-pro–B (B220+CD43+CD19−IgM−), pro-B (B220+CD43+CD19+IgM−), pre-B (B220+CD43−CD19+IgM−), and immature B cells (B220+CD43−CD19+IgM+) isolated from the BM of transplanted mice. Combined data from 5 independent experiments are shown. *P < .05, ***P < .001, ****P < .0001 (Mann-Whitney U test); n = 24 (16 for overall number of cells) and n = 28 (18 for overall number of cells) for the BM only and BM plus T cells groups, respectively. (D) Expression of Ebf1 in sorted pre-pro–B and pro-B progenitors isolated from the BM-only control group (open bars) and cGVHD group (filled bars). Pooled data from 2 independent experiments are shown. ****P < .0001 (unpaired Student t test); n = 6 for each group; sorting purity was >95% and >90% for pre-pro–B and pro-B progenitors, respectively. (E) Expression of Pax5 gene in sorted pro-B and pre-B progenitors isolated from the BM-only control group (open bars) and cGVHD group (filled bars); n = 3 for each group, >90% and >95% for pro-B and pre-B progenitors. Bars represent standard error of the mean. NS, not significant.
Expression of the key transcription factors in donor-derived B-cell progenitors
Early stages of commitment to the B-cell fate are orchestrated by transcription factors involved in the upregulation of B-cell–specific genes and suppression of alternative hematopoietic lineage differentiation, which includes E2A, EBF1, PAX5, and Ikaros. EBF1 is required for B-cell commitment, pro–B-cell development, and subsequent transition to the pre–B-cell stage.21-27 Given the differences we found in the number of pre-pro– vs pro-B cells, we evaluated if this was associated with changes in the expression of Ebf1 and Pax5 using RT-PCR. We found that Ebf1 is significantly downregulated in the pre-pro–B-cell fraction, but not in the more mature pro–B-cell fraction (Figure 3D), in mice receiving T cells compared with BM alone. In contrast, expression of Pax5 in committed B-cell progenitors was not significantly different between the 2 groups (Figure 3E). Taken together, these findings indicate that B lymphoid development is impaired at the earliest stages of commitment in recipients during cGVHD.
Previous work has indicated that BM is one the target organs in aGVHD and that development of B-cell progenitors in mice with aGVHD is impaired. One hypothesis for our findings is that the BO model of cGVHD is characterized by early development of aGVHD. To determine this, we compared cGVHD animals with mice that developed aGVHD as a result of receiving a higher dose (5 × 105 splenic T cells/mouse) of T cells after increased dose of irradiation (750 cGy vs 700 cGy). As expected, mice in the aGVHD group had a significantly higher pathology scores and weight loss compared with the animals in the cGVHD group (supplemental Figure 3). The degree of inflammation in the gastrointestinal tract was significantly different in models of acute versus chronic GVHD. Thus, we evaluated the generation of pro-inflammatory cytokines and tissue pathology in the colon of recipients post-transplant. There was a significant difference in the scoring of GVHD at day 17 in mice with acute compared with chronic GVHD. This correlated with substantially reduced amounts of the proinflammatory cytokines IFN-γ and TNF-α in the colon, which were not substantially different compared with those of mice receiving BM alone in mice with cGVHD (Figure 4A-B). These data indicate that there is little evidence of the proinflammatory environment found in mice with aGVHD using the BO model of cGVHD. Therefore, given the lack of evidence of significant disease and clinical scoring at day 17 posttransplant, the loss of B-cell precursors in the BM is unlikely due to aGVHD.
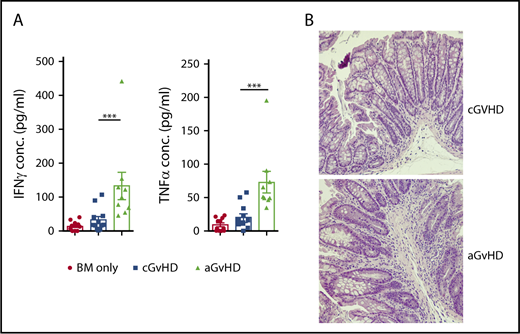
Comparison of cGVHD model and high T-cell dose B6->B10.BR model that mimics aGVHD. B10.BR mice were transplanted with BM only or BM and either a low (5 × 104) or high (5 × 105) dose of T cells from B6 donor mice. Tissues from the animals were harvested on day 17. (A) Levels of TNF-α and IFN-γ in the small intestine of the mice 17 days posttransplant. ***P < .001 (Mann-Whitney U test); n = 12 and n = 9 for cGVHD and aGVHD groups, respectively. Bars represent standard error of the mean. (B) Lymphocyte infiltration in the submucosa of the small intestine of cGVHD (top panel) and aGVHD (bottom panel) animals (hematoxylin and eosin staining, bright-field microscopy using a Nikon Eclipse Ti microscope, original magnification ×100).
Comparison of cGVHD model and high T-cell dose B6->B10.BR model that mimics aGVHD. B10.BR mice were transplanted with BM only or BM and either a low (5 × 104) or high (5 × 105) dose of T cells from B6 donor mice. Tissues from the animals were harvested on day 17. (A) Levels of TNF-α and IFN-γ in the small intestine of the mice 17 days posttransplant. ***P < .001 (Mann-Whitney U test); n = 12 and n = 9 for cGVHD and aGVHD groups, respectively. Bars represent standard error of the mean. (B) Lymphocyte infiltration in the submucosa of the small intestine of cGVHD (top panel) and aGVHD (bottom panel) animals (hematoxylin and eosin staining, bright-field microscopy using a Nikon Eclipse Ti microscope, original magnification ×100).
Donor T cells are critical to impaired B-cell development
Infiltration of alloreactive CD4+ T cells has been shown to disrupt hematopoietic niches in the BM. Osteoblasts form a niche that is important for the support of a subset of hematopoietic stem cells and early B-cell progenitors.28-31 These cells are the target of alloreactive CD4+ T cells in mouse models and patients with aGVHD.14,15,32,33 However, a similar role for donor T cells in the development of BM B cells has not been demonstrated in a murine model of cGVHD. Thus, we evaluated the role that donor T cells play in the impaired development of B cells.
Despite significantly lower total number of donor T cells (5 × 104) used for infusion in the B6→B10.BR model compared with the established aGVHD mouse model (∼100-fold higher), we detected a significant increase in CD4+ donor T cells in the BM of mice with cGVHD compared with control animals (Figure 5A). The increase in donor T cells in the BM was correlated with a decrease in osteoblasts isolated from the tibiae and femurs of mice receiving donor T cells compared with TCD BM alone (Figure 5B). To determine the function that donor T cells play in impaired BM B-cell development, we depleted CD4+ cells in vivo by injecting 2 doses of anti-CD4 monoclonal antibody (mAb) during the first 2 weeks posttransplant (Figure 5A). Depletion of CD4+ T cells (Figure 5A) resulted in an improvement in the frequency of osteoblasts compared with control antibody treatment (Figure 5B). To further examine the effects of CD4+ T-cell depletion, we evaluated B-cell development in the BM in mice receiving anti-CD4 mAb compared with control. A significant improvement in the frequency and overall number of pro-B, pre-B, and immature B cells was found after depletion of CD4+ T cells (Figure 5C).
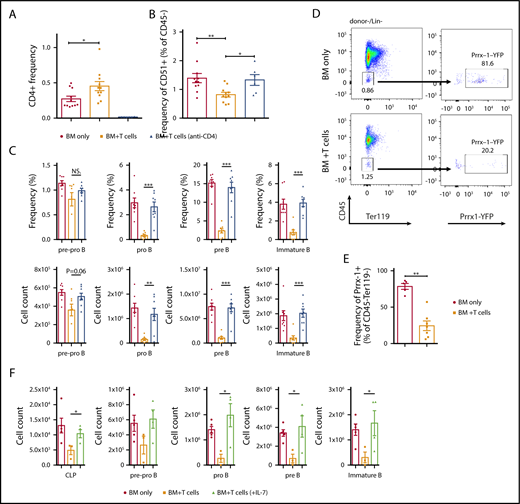
CD4+T-cell–dependent aberration of B-cell progenitor niche in cGVHD animals. B10.BR mice were transplanted with BM only or BM and T cells from B6 donor mice. A subgroup of BM plus T-cell animals received injections of anti-CD4 on days 5, 7, 12, and 14 posttransplantation (GK1.5, 0.5 mg/mouse). Tibiae and femurs for osteoblast analysis and BM were harvested on day 30. (A) Frequency of infiltrating CD4+ T cells in the BM on day 30. Combined data from 2 independent experiments. *P < .05 (Mann-Whitney U test); n = 11 for BM only and BM plus T cells, and n = 6 for BM+T (anti-CD4). (B) Frequency of osteoblasts isolated from tibiae and femurs (percentage of Ter119−CD31−CD51+cells in the CD45− gate) 30 days posttransplantation. Combined data from 2 independent experiments, *P < .05, **P < .01 (Mann-Whitney U test), N = 11 for BM only and BM+T cells, N = 6 for BM+T (anti-CD4). (C) Frequency determined from the total number of cells in the BM (top) and overall number (bottom) of donor derived pre-pro B (B220+CD43+CD19−IgM−), pro B (B220+CD43+CD19+IgM−), pre B (B220+CD43-CD19+IgM−) and immature B cells (B220+CD43-CD19+IgM+) isolated from the BM of transplanted mice. Combined data from 2 independent experiments are shown. ***P < .001 (Mann-Whitney U test); n = 8 for all groups. (D-E) Prrx-1+YFP+ mice on a B6 background were transplanted with BM only or BM and T cells from B10.BR donor mice, and representative flow cytometry plots (D) and frequency of Prrx-1 YFP+ perivascular cells isolated from tibiae and femurs in CD45-Ter119− gate at day 30 after the transplantation (E) are shown. Combined data from 3 independent experiments are shown. **P < .01 (Mann-Whitney U test); n = 5 for BM only and n = 7 for BM plus T cells. (F) Overall number of donor-derived CLP, pre-pro–B (B220+CD43+CD19−IgM−), pro-B (B220+CD43+CD19+IgM−), pre-B (B220+CD43−CD19+IgM−), and immature B cells (B220+CD43−CD19+IgM+) isolated from the BM of transplanted mice. The BM plus T group received daily injections of either saline (black bars) or 1 μg/mouse mIL-7 (green bars) during the first 2 weeks posttransplantation. *P < .05 (Mann-Whitney U test); n = 5 for BM only, n = 3 for BM plus T cells (saline), and n = 4 for BM plus T cells (mIL-7). Bars represent standard error of the mean.
CD4+T-cell–dependent aberration of B-cell progenitor niche in cGVHD animals. B10.BR mice were transplanted with BM only or BM and T cells from B6 donor mice. A subgroup of BM plus T-cell animals received injections of anti-CD4 on days 5, 7, 12, and 14 posttransplantation (GK1.5, 0.5 mg/mouse). Tibiae and femurs for osteoblast analysis and BM were harvested on day 30. (A) Frequency of infiltrating CD4+ T cells in the BM on day 30. Combined data from 2 independent experiments. *P < .05 (Mann-Whitney U test); n = 11 for BM only and BM plus T cells, and n = 6 for BM+T (anti-CD4). (B) Frequency of osteoblasts isolated from tibiae and femurs (percentage of Ter119−CD31−CD51+cells in the CD45− gate) 30 days posttransplantation. Combined data from 2 independent experiments, *P < .05, **P < .01 (Mann-Whitney U test), N = 11 for BM only and BM+T cells, N = 6 for BM+T (anti-CD4). (C) Frequency determined from the total number of cells in the BM (top) and overall number (bottom) of donor derived pre-pro B (B220+CD43+CD19−IgM−), pro B (B220+CD43+CD19+IgM−), pre B (B220+CD43-CD19+IgM−) and immature B cells (B220+CD43-CD19+IgM+) isolated from the BM of transplanted mice. Combined data from 2 independent experiments are shown. ***P < .001 (Mann-Whitney U test); n = 8 for all groups. (D-E) Prrx-1+YFP+ mice on a B6 background were transplanted with BM only or BM and T cells from B10.BR donor mice, and representative flow cytometry plots (D) and frequency of Prrx-1 YFP+ perivascular cells isolated from tibiae and femurs in CD45-Ter119− gate at day 30 after the transplantation (E) are shown. Combined data from 3 independent experiments are shown. **P < .01 (Mann-Whitney U test); n = 5 for BM only and n = 7 for BM plus T cells. (F) Overall number of donor-derived CLP, pre-pro–B (B220+CD43+CD19−IgM−), pro-B (B220+CD43+CD19+IgM−), pre-B (B220+CD43−CD19+IgM−), and immature B cells (B220+CD43−CD19+IgM+) isolated from the BM of transplanted mice. The BM plus T group received daily injections of either saline (black bars) or 1 μg/mouse mIL-7 (green bars) during the first 2 weeks posttransplantation. *P < .05 (Mann-Whitney U test); n = 5 for BM only, n = 3 for BM plus T cells (saline), and n = 4 for BM plus T cells (mIL-7). Bars represent standard error of the mean.
In addition to osteoblasts, several groups have found that Prrx1+ perivascular stromal cells are a critical component of the niche for hematopoietic stem cells, CLP cells, and early B-cell progenitors.28,29 To evaluate whether cGVHD also targeted this subset of perivascular stromal cells, we used Prrx1 conditional mice in which Prrx1-expressing cells express YFP.29,34 We observed a decrease of Prrx1-expressing perivascular stromal cells in the animals that received donor T cells compared with the TCD BM alone (Figure 5D-E).
One of the critical functions of osteoblasts and perivascular stromal cells is production of cytokines, such as interleukin-7 (IL-7), and chemokines, such as CXCL12. To determine whether IL-7 supplementation can rescue BM B-cell development in the mouse model of cGVHD, we administered IL-7 to the animals that received donor T cells along with TCD BM. IL-7 injections had no effect on numbers of Flt3+ and Flt3− LSK cells but resulted in an increase of the CLP and committed pro-B, pre-B, and immature B-cell progenitors in the BM compared with the group that received saline injections (Figure 5F; supplemental Figure 4).
Thus, cGVHD is associated with a significant decrease in both osteoblasts and perivascular stromal cells, which are critical for B-cell development in the BM. This process was mediated in part by donor CD4+ T cells. This effect can be ameliorated by the external supplementation of IL-7, which we hypothesize compensated for the absence of this cytokine in the BM.
In vivo anti-DR3 expanded Tregs reduce organ pathology and partially restore lung function in a cGVHD model
Previous reports have suggested that immune tolerance may exist in the BM niche secondary to the presence of Tregs.35,36 Activation and proliferation of Tcons in an aGVHD model can be prevented by in vivo expansion of potent Tregs using activation of TNF superfamily receptor TNFRSF25 (also termed DR3). Expansion of donor Tregs can be achieved by injection of either a single dose of agonistic anti-DR3 mAb (4C12)37,38 or TL1A-Ig fusion protein along with low-dose IL-2.39,40 To determine whether Tregs can protect the components of BM niche during B-cell development in mice that develop cGVHD, we treated T-cell donor mice with anti-DR3 mAb before transplantation.
As previously reported, anti-DR3 mAb injection increased the frequency of splenic CD4+FoxP3GFP+ cells in the donor animals37 (Figure 6A). Mice infused with the same number of total T cells from anti-DR3–treated donors had lower pathology scores and weight loss compared with animals in cGVHD group that received T cell isolated from isotype-treated donors (supplemental Figure 5A-B). We next performed histological analysis of cGVHD target organs from mice receiving TCD BM plus donor T cells with anti-DR3 mAb compared with those receiving the same transplant with irrelevant control mAb. Perivascular collagen deposition in the parenchymal part of the liver and infiltration of lymphocytes characteristic of cGVHD was diminished in the anti-DR3 group compared with the isotype-control group (Figure 6B). We observed a decrease in pathological changes in the lung, including formation of collagen around the bronchioles and peribronchiolar lymphocyte infiltration (Figure 6C). Treatment of donor animals with anti-DR3 antibody prior to transplantation also improved respiratory function in cGVHD model compared with isotype-treated donors, as evidenced by higher arterial oxygen saturation measured by pulse oximetry (Figure 6D).
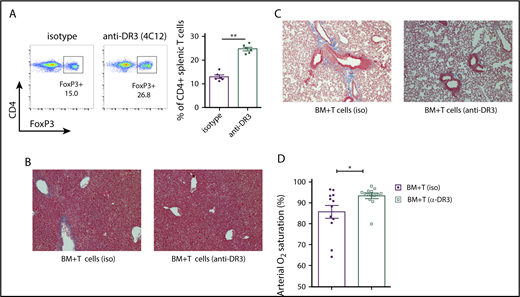
Infusion of splenic T cells from anti-DR3–treated donors reduces cGVHD severity. (A) Representative flow cytometry plot and a summary of 6 independent experiments indicating expansion of splenic FoxP3-GFP+ donor cells in the CD4+ population after a single dose of 0.5 mg/kg anti-DR3 antibody (4C12) 3 days before the transplantation. **P < .01. (B-C) Collagen deposition in perivascular regions of liver parenchyma (B; blue staining) and the peribronchiolar area of the lung (C; Masson’s trichrome staining, bright-field microscopy using Nikon Eclipse Ti microscope, original magnification ×200). (D) Arterial oxygen saturation (SpO2) measured using pulse oximetry in cGVHD (iso) and cGVHD (anti-DR3) animals. *P < .05 (Mann-Whitney U test); n = 12 for cGVHD (iso) and n = 14 for cGVHD (anti-DR3). Bars represent standard error of the mean.
Infusion of splenic T cells from anti-DR3–treated donors reduces cGVHD severity. (A) Representative flow cytometry plot and a summary of 6 independent experiments indicating expansion of splenic FoxP3-GFP+ donor cells in the CD4+ population after a single dose of 0.5 mg/kg anti-DR3 antibody (4C12) 3 days before the transplantation. **P < .01. (B-C) Collagen deposition in perivascular regions of liver parenchyma (B; blue staining) and the peribronchiolar area of the lung (C; Masson’s trichrome staining, bright-field microscopy using Nikon Eclipse Ti microscope, original magnification ×200). (D) Arterial oxygen saturation (SpO2) measured using pulse oximetry in cGVHD (iso) and cGVHD (anti-DR3) animals. *P < .05 (Mann-Whitney U test); n = 12 for cGVHD (iso) and n = 14 for cGVHD (anti-DR3). Bars represent standard error of the mean.
Expansion of Tregs after treatment with anti-DR3 mAb resulted in a modest reduction of CD4+FoxP3− cells in the T-cell inoculum that could contribute to the observed effects on cGVHD pathology. To rule out this possibility, we coadministered total splenic donor T cells with 2 × 104 donor Tregs (∼2.5:1 ratio of Tcons to Tregs). Infusion of this very modest number of Tregs was sufficient to significantly improve the recovery of pro-B and pre-B cells and showed a trend toward improvement of immature B cells (supplemental Figure 6). These data demonstrate that the beneficial effects of the administration of donor Tregs in B-cell development and strongly argue against the benefit of anti-DR3 mAb therapy being mediated by the very modest decrease in donor CD4+ Tcons.
Infusion of T cells from anti-DR3–treated donors protects endosteal osteoblasts and improves BM B-cell development in cGVHD
To determine whether the beneficial effect of expanded donor Tregs also can affect B-cell development, we evaluated B-cell development in mice receiving donor splenic T cells from anti-DR3–treated donors. We found an increase in the frequency and overall number of CLP cells in the BM of animals receiving anti-DR3–treated donor cells compared with those receiving isotype antibody (Figure 7A). We also observed significantly higher frequency and total number of pro-B, pre-B, and immature B cells in the BM of cGVHD mice infused with T cells from anti-DR3–treated donors (Figure 7B-D). Interestingly, the overrepresentation of pre-pro–B cells detected in the B220+ fraction of BM in cGVHD animals was also significantly reduced in mice receiving donor T cells after anti-DR3 treatment (7.91% ± 0.62 vs 14.64% ± 2.75).
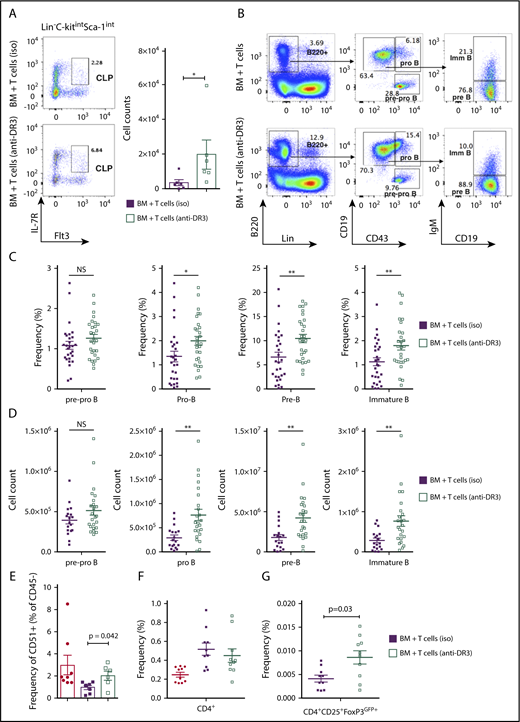
Transplantation of low-dose of splenic T cells from anti-DR3–pretreated donors protected osteoblasts and partially rescued impaired BM B-cell development in cGVHD. B10.BR mice were transplanted with BM only or BM and T cells from B6 donor mice injected with either anti-DR3 mAb (4C12, 0.5 mg/kg) or isotype control IgG (NTK888) 3 days prior to the procedure. (A) Representative flow cytometry plots demonstrating frequency of donor-derived CLP cells in Lin−c-KitintSca-1int and total number of donor-derived CLP cells. Combined data from 2 independent experiments are shown. *P < .05, **P < .01 (Mann-Whitney U test); n = 6 in each group. (B) Representative flow cytometry plots demonstrating frequency of pre-pro–B, pro-B, pre-B, and immature B cells in the BM (numbers indicates frequency of parental gate). (C-D) Frequency of total BM (C) and total number (D) of donor-derived pre-pro–B (B220+CD43+CD19−IgM−), pro B (B220+CD43+CD19+IgM−), pre B (B220+CD43−CD19+IgM−) and immature B cells (B220+CD43−CD19+IgM+) isolated from BM of transplanted mice. Combined data from 4 independent experiments are shown. *P < .05; **P < .01 (Mann-Whitney U test); n = 18 and n = 24 for BM plus T cells (iso) and BM plus T cells (anti-DR3), respectively. (E) Frequency of osteoblast isolated from tibiae and femurs (percentage of Ter119−CD31−CD51+cells in the CD45− gate). Combined data from 2 independent experiments are shown. Mann-Whitney U test; n = 8 for BM only and BM plus T cells, and n = 6 for BM plus T cells (anti-DR3). (F-G) Frequency of infiltrating donor-derived CD4+ T cells (F) and CD4+CD25+FoxP3GFP+ Tregs (G) in the BM. The BM-only group was not included in this analysis due to lack donor-derived FoxP3-GFP–expressing Tregs. Combined data from 3 independent experiments are shown. Mann-Whitney U test; n = 10 for all groups. Bars represent standard error of the mean.
Transplantation of low-dose of splenic T cells from anti-DR3–pretreated donors protected osteoblasts and partially rescued impaired BM B-cell development in cGVHD. B10.BR mice were transplanted with BM only or BM and T cells from B6 donor mice injected with either anti-DR3 mAb (4C12, 0.5 mg/kg) or isotype control IgG (NTK888) 3 days prior to the procedure. (A) Representative flow cytometry plots demonstrating frequency of donor-derived CLP cells in Lin−c-KitintSca-1int and total number of donor-derived CLP cells. Combined data from 2 independent experiments are shown. *P < .05, **P < .01 (Mann-Whitney U test); n = 6 in each group. (B) Representative flow cytometry plots demonstrating frequency of pre-pro–B, pro-B, pre-B, and immature B cells in the BM (numbers indicates frequency of parental gate). (C-D) Frequency of total BM (C) and total number (D) of donor-derived pre-pro–B (B220+CD43+CD19−IgM−), pro B (B220+CD43+CD19+IgM−), pre B (B220+CD43−CD19+IgM−) and immature B cells (B220+CD43−CD19+IgM+) isolated from BM of transplanted mice. Combined data from 4 independent experiments are shown. *P < .05; **P < .01 (Mann-Whitney U test); n = 18 and n = 24 for BM plus T cells (iso) and BM plus T cells (anti-DR3), respectively. (E) Frequency of osteoblast isolated from tibiae and femurs (percentage of Ter119−CD31−CD51+cells in the CD45− gate). Combined data from 2 independent experiments are shown. Mann-Whitney U test; n = 8 for BM only and BM plus T cells, and n = 6 for BM plus T cells (anti-DR3). (F-G) Frequency of infiltrating donor-derived CD4+ T cells (F) and CD4+CD25+FoxP3GFP+ Tregs (G) in the BM. The BM-only group was not included in this analysis due to lack donor-derived FoxP3-GFP–expressing Tregs. Combined data from 3 independent experiments are shown. Mann-Whitney U test; n = 10 for all groups. Bars represent standard error of the mean.
We next evaluated if the improvement in B-cell development after the infusion of anti-DR3–treated splenic T cells was due, in part, to an increased frequency of Tregs. The frequency of Lin−CD45−CD31−CD51+ osteoblasts was higher in the recipient mice receiving anti-DR3–treated splenocytes than in animals in the control group (Figure 7E). This effect was not mediated by changes in the frequency of donor CD4+ Tcons, which were not changed in mice receiving anti-DR3 mAb versus control (Figure 7F). However, the frequency of CD4+CD25+FoxP3GFP+ cells in BM of mice receiving anti-DR3–treated T cells was significantly greater 30 days after transplantation (Figure 7G). Taken together these data indicate that the presence of anti-DR3 expanded CD4+CD25+FoxP3GFP+ Tregs in the BM results in a higher Treg-to-Tcon ratio, which was associated with a protective effect on osteoblasts that form the BM niche for early B-cell progenitors. The increased number of osteoblasts correlated with improved recovery of BM B-cell development in the B6→B10.BR model of cGVHD.
Discussion
cGVHD remains a significant cause of morbidity and mortality in patients undergoing allo-SCT. The B-cell repertoire in patients with cGVHD is altered with diminished generation of antigen-specific antibody often in the setting of B-cell lymphopenia.9,10 The mechanism for the diminished generation of antigen-specific antibody with increased production of auto-/alloreactive antibodies in patients with cGVHD is not known. Here, we demonstrated that B-cell development in the BM is impaired in the BO model of cGVHD. This was associated with donor T-cell–mediated loss of cells critical for the generation of BM B cells. Interestingly, we found that donor Tregs were capable, in part, of restoring the development of B cells in the BM, which was associated with diminished target organ damage as a result of cGVHD. Additionally, we found that the effects of donor T cells on the BM compartment had a differential effect on specific B-cell precursors. The frequency and absolute number of pre-, pro-, and immature B cells were significantly lower in mice that developed cGVHD. The reduction in the more primitive pre-pro–B progenitor population, however, was more moderate.
During the process of differentiation from CLPs to the immature B-cell stage, progenitors occupy several BM niches. During this process, there is a network of stage-specific transcription factors (E2A family, EBF1, PAX5, and others) that facilitate commitment to B-cell fate and suppress alternative lineage differentiation.30 Here, aberrations in the development of lineage-committed B-cell progenitors were manifested at the molecular level by lower levels of expression of the key transcription factor Ebf1 at the pre-pro–B to pro–B-stage transition. We hypothesize that the modest reduction in total number of pre-pro–B cells that we observed was due to the low requirements for Ebf1 expression at this stage and ability of these cells to develop in an IL-7–independent fashion.41,42 Consistent with our analysis of B-cell progenitors in the BM, there was severe impairment in the transition of pre-pro–B cells with low Ebf1 expression to the pro-B stage.42 Ebf1 has been also shown to be regulated via IL-7–independent pathways in committed B-cell progenitors, which can in part explain the higher levels of Ebf1 in more mature populations.43
Previous work has found impaired B-cell development in mice with aGVHD. One hypothesis for our findings was that the impaired development of B cells in the BO model of cGVHD is due to early onset of aGVHD. We believe that our findings are not consistent with this interpretation. First, we did not observe a statistically significant decrease in the total number of LSK cells, which has been reported in the study demonstrating impaired B-cell development in mice with aGVHD.13 Second, we found no evidence of gastrointestinal tract tissue damage mediated by acute GVHD at the time of impaired B-cell development in the BM. Finally, our clinical scoring at day 17 was inconsistent with significant evidence of acute GVHD. Our findings that the development of cGVHD did not impact the number of LSK cells strongly suggest that the reduction of lymphoid-lineage B-cell progenitors was not due to a decreased number of cells that give rise to CLP cells.
Previous work suggested that Tregs traffic to the BM partly through CXCR4/CXCL12 interactions,44 and recent work indicates that presence of Tregs may be critical to the steady-state generation of B cells.36 We found that infusion of total splenic T cells from anti-DR3–treated donors that had a higher frequency of Tregs had a protective effect on B-cell development in cGVHD animals. Infusion of anti-DR3–treated splenic T cells was also effective in enhancing BM B-cell development in recipients infused with allogeneic T and BM cells. We chose to focus on a strategy that used expansion of donor-derived Treg cells, as this method preserved the graft-versus-tumor effect associated with BMT and, therefore, would be more clinically relevant.37,39 The reduced pathology in cGVHD animals after the receipt of donor T cells with increased numbers of Tregs is likely a combination of the early effects (improvement of BM B-cell development) and late effects (GC reaction inhibition in spleen),40 as well as potential inhibitory activity of Tregs in other tissues. In addition, other components of the immune system (eg, CD8+ T cells) can be affected by expansion of Tregs and therefore contribute to the reduction of pathological manifestation of cGVHD and potentially the improvement in B-cell progenitors in the BM.
Patients with steroid-refractory cGVHD that received low-dose IL-2 had significant improvement, with an overall response rate of 52%.45 This correlated with a marked significant increase in Tregs in the bloodstream. There was no association in that study with infusion of IL-2 and the number of B cells, although the investigators did not specifically evaluate for the presence of naive B cells. A second evaluation demonstrated that infusion of IL-2 led to increased proliferation, increased thymic export, and enhanced Treg survival during the infusion of IL-2.46 One potential hypothesis for the activity of low-dose IL-2 is the ability to improve B-cell development in the BM, which would lead to an enhanced number of naive B cells exported to the peripheral compartment. By outcompeting low-affinity alloreactive B cells for IL-7 and B-cell activating factor, the increased number of naive B cells would diminish the generation of antibodies that may be critical to the pathogenesis of cGVHD. This hypothesis can be evaluated in future studies of patients receiving low-dose IL-2 therapy.
In summary, we have demonstrated that B-cell development in the BM is disrupted in cGVHD, which correlated with donor T-cell infiltration into the BM in recipient mice. Infusion of T cells with an enhanced number of Tregs was effective in reversing the effects of donor T cells on B-cell development in the BM. Thus, enhancing the number of donor Tregs may provide a mechanism for diminishing cGVHD in patients by enhancing the development of B-cell precursors in the BM.
The full-text version of this article contains a data supplement.
Acknowledgments
The authors thank the UNC Flow Cytometry Core for performing fluorescence-activated cell sorting.
The UNC Flow Cytometry Core Facility is supported in part by the National Institutes of Health (NIH), National Cancer Institute (P30 CA016086; Cancer Center Core Support Grant to the UNC Lineberger Comprehensive Cancer Center) and North Carolina Biotech Center Institutional Support Grant 2012-IDG-1006. Animal histopathology was performed in the LCCC Animal Histopathology Core Facility at the University of North Carolina at Chapel Hill. The LCCC Animal Histopathology Core is supported in part by an NIH, National Cancer Institute Center Core Support Grant (2P30CA016086-40) to the UNC Lineberger Comprehensive Cancer Center. This work was supported by the NIH, National Heart, Lung, and Blood Institute (R01 HL139730 [J.S.S.], R01 HL11879 [B.R.B.], and R01 HL129061 [S.S.]) and National Cancer Institute (R01 CA166794 [J.S.S.] and P01 CA142106 [B.R.B.]).
Authorship
Contribution: O.V.K. performed experiments and wrote the manuscript; T.A.D., H.B., and D.W.B. performed experiments and edited the manuscript; K.A.F. performed experiments; K.P.M. and S.T. edited the manuscript; S.S., B.R.B., and J.M.C. reviewed and edited the manuscript; and J.S.S. conceived of and supervised completion of this project and reviewed and edited the manuscript.
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Correspondence: Jonathan S. Serody, University of North Carolina at Chapel Hill, 5012 Marsico Hall, 125 Mason Farm Rd, Chapel Hill, NC 27599-7295; e-mail: jonathan_serody@med.unc.edu.