

Key Points
Fine-tuning of chromatin accessibility primes HSPC transition from nascent to fetal states.
Smarca5 interacts with nucleolin to promote chromatin remodeling and facilitate genomic binding of hematopoietic TFs in fetal HSPCs.

Abstract
Nascent hematopoietic stem and progenitor cells (HSPCs) acquire definitive hematopoietic characteristics only when they develop into fetal HSPCs; however, the mechanisms underlying fetal HSPC development are poorly understood. Here, we profiled the chromatin accessibility and transcriptional features of zebrafish nascent and fetal HSPCs using ATAC-seq and RNA-seq and revealed dynamic changes during HSPC transition. Functional assays demonstrated that chromatin remodeler-mediated epigenetic programming facilitates fetal HSPC development in vertebrates. Systematical screening of chromatin remodeler-related genes identified that smarca5 is responsible for the maintenance of chromatin accessibility at promoters of hematopoiesis-related genes in fetal HSPCs. Mechanistically, Smarca5 interacts with nucleolin to promote chromatin remodeling, thereby facilitating genomic binding of transcription factors to regulate expression of hematopoietic regulators such as bcl11ab. Our results unravel a new role of epigenetic regulation and reveal that Smarca5-mediated epigenetic programming is responsible for fetal HSPC development, which will provide new insights into the generation of functional HSPCs both in vivo and in vitro.
Introduction
The hematopoietic lineage tree is rooted from the hematopoietic stem and progenitor cells (HSPCs), which are capable of self-renewal and multilineage differentiation.1-3 In vertebrates, the earliest definitive HSPCs (ie, nascent HSPCs) arise from the ventral wall of dorsal aorta in the aorta-gonad-mesonephros (AGM) region, through the endothelial-to-hematopoietic transition.4-6 Then, nascent HSPCs migrate into a transient fetal hematopoietic organ, fetal liver (FL) in mammals or caudal hematopoietic tissue (CHT) in zebrafish, for rapid expansion/differentiation (ie, fetal HSPCs).7,8 Previous studies in mice have revealed that most nascent pre-HSCs are immature, and these cells acquire the complete transcriptome signature of definitive HSCs only when they colonize the FL.9,10 Moreover, transplantation assays have shown that nascent HSPCs, susceptible to immune-rejection, preferentially engraft mouse neonates for effective transplant outcomes.11 Given that there are substantial molecular and phenotypical differences between nascent and fetal HSPCs, understanding the intrinsic mechanism facilitating fetal HSPC development can help improve the strategy to generate functional HSPCs in vitro.
The programming of epigenome is known to contribute to the stepwise cellular development.12,13 Among the complicated epigenetic regulation, chromatin accessibility is critical for gene expression, as the nucleosome-free DNA is accessible to transcription factors (TFs).14 The ATPase-dependent chromatin remodelers, including switch/sucrose nonfermentable (SWI/SNF), imitation SWI (ISWI), chromodomain helicase DNA-binding (CHD), and inositol requiring 80 (INO80), use the energy of ATP hydrolysis to participate in noncovalent ordering of chromatin structure.15-17 Chromatin remodelers play an essential role in hematopoietic development. For instance, CHD1 is essential for HSPC emergence from hemogenic endothelial cells in the AGM region by upregulating the overall nascent transcription in mice.18 Brg1, a member of SWI/SNF family, plays an important role in B-cell development by facilitating access of B-lineage regulators to enhancer repertoires.19 However, whether and how chromatin remodelers are involved in fetal HSPC development by mediating chromatin accessibility remain elusive.
Here, we profiled the chromatin accessibility landscape and transcriptome feature by performing assay for transposase-accessible chromatin with high-throughput sequencing (ATAC-seq) and RNA sequencing (RNA-seq) analyses in nascent and fetal HSPCs of zebrafish. Systematical screening identified a chromatin remodeler smarca5, and loss of smarca5 led to inaccessible chromatin regions in genes involved in HSPC expansion and differentiation, thereby impairing fetal HSPCs. We further identified a coremodeler, nucleolin (Ncl), which collaborates with Smarca5 to facilitate chromatin remodeling for hematopoietic TF binding. Our study provides a comprehensive understanding of nascent-to-fetal HSPC transition and uncovers a pivotal role of epigenetic programming in fetal HSPC development.
Methods
Zebrafish and mouse strains
Zebrafish strains including Tubingen, Tg (CD41:GFP),20 Tg (gata1:dsRed),21 Tg (kdrl:mCherry),4 Tg (mpo:GFP),22 Tg (lyz:dsRed),23 Tg (coro1a:GFP),24 p53M214K,25 smarca5zko1049a heterozygous mutants, Tg (hsp70:flag-smarca5-EGFP), Tg (hsp70:flag-nucleolin-EGFP), and nucleolinm4 heterozygous mutants were raised under standard conditions (28.5°C in system water). The zebrafish embryos were raised in an incubator at 28.5°C. Wild-type C57BL/6 mice were purchased from SPF (Beijing) Biotechnology Co Ltd. The morning of detection of vaginal plug was defined as E0. This study was approved by the Ethical Review Committee in the Institute of Zoology, Chinese Academy of Sciences, China.
Procedures and methods
Detailed information about whole-mount in situ hybridization (WISH), Morpholinos, generation of smarca5 and ncl mutants using CRISPR/Cas9, western blotting, coimmunoprecipitation (CoIP), RNA-seq, ATAC-seq, chromatin immunoprecipitation sequencing (ChIP-seq), and the following bioinformatics analysis is provided in the supplemental Methods, available on the Blood Web site.
Image analysis
Raw image data were processed using ImageJ, photoshop CC 2018, and Adobe Illustrator CC 2018.
Statistical analysis
All statistical analysis was performed for at least 3 independent biological repeats. GraphPad Prism 6 was used to analyze the data. Data are mean ± standard deviation (SD). P values calculated by 2-tailed, unpaired Student t test were used to indicate the significance if not clarified in figure legends.
Results
Dynamic chromatin accessibility underlies transcriptional program differences between nascent and fetal HSPCs
The molecular and phenotypical differences between nascent and fetal HSPCs prompt us to explore the intricate mechanism driving the formation of mature definitive HSPCs during nascent-to-fetal HSPC transition. Chromatin accessibility is critical for gene expression, as the accessible sites of DNA are usually bound by TFs to turn on or off transcription. To investigate the effects of chromatin accessibility on the temporally specific feature of HSPCs, ATAC-seq was performed for nascent and fetal HSPCs (CD41:GFP+gata1:dsRed−) (Figure 1A).20,26 Pearson correlation analysis indicated high reproducibility of ATAC-seq data after normalization (supplemental Figure 1A). By profiling accessible regions at promoters (TSS ± 2 kb) and distal regulatory regions (outside promoter) (supplemental Figure 1B), we found that despite the presence of a large number of peaks at conserved accessible promoters (“both accessible” [9624]), nascent and fetal HSPCs are characterized by stage-specific accessible regions (“nascent HSPC-specific accessible” [1324] and “fetal HSPC-specific accessible” [467]). Moreover, such dynamic changes in chromatin accessibility were also observed in distal regulatory regions (“nascent HSPC-specific accessible” [10 963], “both accessible” [8101], and “fetal HSPC-specific accessible” [2230]), suggesting the dynamic change in chromatin accessibility in nascent HSPCs transitioning into fetal HSPCs.
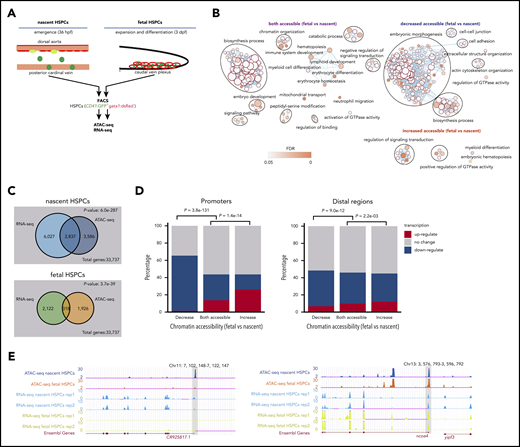
The chromatin accessibility and transcriptome comparison between nascent and fetal HSPCs. (A) Schematic representation of nascent HSPCs at 36 hours postfertilization (hpf) in the AGM region and fetal HSPCs at 3 days postfertilization (dpf) in the CHT region and the experimental flow to profile accessible chromatin and transcriptome in nascent and fetal HSPCs by ATAC-seq and RNA-seq. (B) GO analysis for genes with both accessible (left), decreased accessible in fetal HSPC (ie, nascent HSPC-specific accessible) (top right), and increased accessible in fetal HSPC (ie, fetal HSPC-specific accessible) (bottom right) chromatin regions. Enrichment scores were calculated by g:Profiler, and the enrichment network was performed by EnrichmentMap in Cytoscape. (C) Venn plot showing the overlap of genes with specific accessible chromatin regions and upregulated expression in nascent or fetal HSPCs, respectively. Genes for ATAC-seq were assigned by all ATAC-seq peak (see supplemental Gene Ontology (GO) analysis section). The P value shows the correlation significance between ATAC-seq and RNA-seq data at these 2 stages. P values were calculated by a 2-tailed χ2 test. (D) Bar plot showing the percentage of “upregulated transcripts,” “no change transcripts,” and “downregulated transcripts” for genes with different states of accessible chromatin at nascent and fetal stages at promoters (left) and distal regulatory regions (right). (E) The genome browser views showing ATAC-seq and RNA-seq signals of genes specifically expressed in nascent HSPCs (left) or both in nascent and fetal HSPCs (right). FACS, fluorescence-activated cell sorter; FDR, false discovery rate.
The chromatin accessibility and transcriptome comparison between nascent and fetal HSPCs. (A) Schematic representation of nascent HSPCs at 36 hours postfertilization (hpf) in the AGM region and fetal HSPCs at 3 days postfertilization (dpf) in the CHT region and the experimental flow to profile accessible chromatin and transcriptome in nascent and fetal HSPCs by ATAC-seq and RNA-seq. (B) GO analysis for genes with both accessible (left), decreased accessible in fetal HSPC (ie, nascent HSPC-specific accessible) (top right), and increased accessible in fetal HSPC (ie, fetal HSPC-specific accessible) (bottom right) chromatin regions. Enrichment scores were calculated by g:Profiler, and the enrichment network was performed by EnrichmentMap in Cytoscape. (C) Venn plot showing the overlap of genes with specific accessible chromatin regions and upregulated expression in nascent or fetal HSPCs, respectively. Genes for ATAC-seq were assigned by all ATAC-seq peak (see supplemental Gene Ontology (GO) analysis section). The P value shows the correlation significance between ATAC-seq and RNA-seq data at these 2 stages. P values were calculated by a 2-tailed χ2 test. (D) Bar plot showing the percentage of “upregulated transcripts,” “no change transcripts,” and “downregulated transcripts” for genes with different states of accessible chromatin at nascent and fetal stages at promoters (left) and distal regulatory regions (right). (E) The genome browser views showing ATAC-seq and RNA-seq signals of genes specifically expressed in nascent HSPCs (left) or both in nascent and fetal HSPCs (right). FACS, fluorescence-activated cell sorter; FDR, false discovery rate.
Enriched functional gene ontology (GO) analysis was performed for genes with nascent HSPC-specific accessible, both accessible, and fetal HSPC-specific accessible promoter regions, respectively (Figure 1B). The results showed that the enrichment in the GO terms of “erythrocyte homeostasis and differentiation,” “myeloid cell differentiation,” and “catabolic process” was commonly observed in nascent and fetal HSPCs, indicating that the chromatin accessibility of hematopoiesis-related genes was maintained during HSPC transition. The accessible chromatin regions of genes related to “cell-cell junction,” “cell adhesion,” and “embryonic morphogenesis” terms were closed in fetal HSPCs, suggesting the loss of partial characteristics of nascent HSPCs, including endothelial identities. Importantly, the percentage of hematopoietic genes with accessible promoter regions in fetal HSPCs was higher than that in nascent HSPCs (supplemental Figure 1C), indicating that fetal HSPCs have acquired specialized hematopoietic properties compared with nascent HSPCs. Consistently, analysis of the transcriptome dynamics of HSPCs using our previous RNA-seq data27 revealed the acquisition of specialized hematopoietic transcriptome characteristics in fetal HSPCs (supplemental Figure 1D), which coincides with the aforementioned ATAC-seq data. Furthermore, analyzing the overlap of genes with specific chromatin-accessible region and genes highly expressed in nascent or fetal HSPCs suggests a strong correlation between chromatin accessibility and gene expression (Figure 1C). The chromatin accessibility at promoters is more correlated with transcriptional dynamics compared with that at the distal regulatory regions (Figure 1D). Representative examples of ATAC-seq peaks of genes specifically expressed in nascent HSPCs or in both nascent and fetal HSPCs were shown, respectively (Figure 1E). Taken together, the dynamic chromatin accessibility at promoters is important for nascent-to-fetal HSPC transition. Specifically, the higher percentage and expression level of hematopoietic genes within accessible chromatin regions in fetal HSPCs imply that nascent HSPCs are acquiring robust definitive characteristics upon transitioning into fetal HSPCs.
Systematical screening of chromatin remodelers important for fetal hematopoiesis
The dynamic chromatin accessibility underlies transcriptional program differences between nascent and fetal HSPCs, raising questions regarding which factors primarily participate in the regulation of chromatin accessibility landscape. Given that ATP-dependent chromatin remodelers are fundamental for chromatin organization and play critical roles in cell-state transition during development,28-30 we further examined whether the chromatin remodelers participate in the regulation of chromatin accessibility landscape in nascent and fetal HSPCs. We first analyzed the transcriptional levels of ATP-dependent chromatin remodelers, including 65 genes from 4 major families and found that the expression level of chromatin remodelers changed as HSPCs develop from nascent to fetal states (Figure 2A). For instance, the expression of pbrm1 and smarcd3a was enriched in nascent HSPCs, whereas enriched expression of smarca5, baz1a, and smarcd2 was detected in fetal HSPCs. To visualize the dynamic expression of chromatin remodelers, we further performed the self-organizing map (SOM) of RNA-seq data in nascent and fetal HSPCs (Figure 2B). Based on the expression pattern and roles of each member in the 4 families, upregulated genes in fetal HSPCs were selected for further analysis. These include smarca5 and bptf from the ISWI family, smarca4a and smarca2 from the SWI/SNF family, chd2 and chd4b from the CHD family, as well as ino80 from the INO80 family (Figure 2B-C).
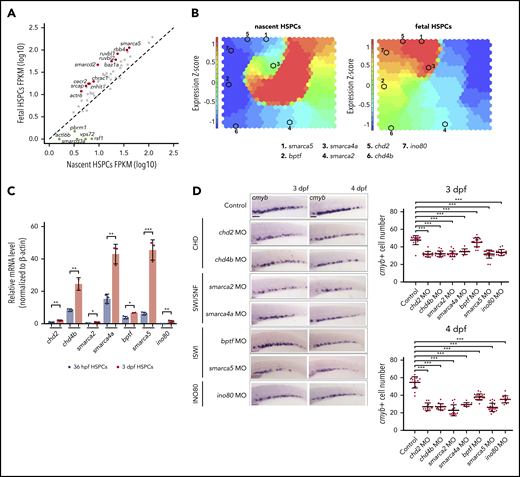
Systematical screening of chromatin remodelers controlling fetal HSPC development. (A) Scatter plots showing the expression level of chromatin remodeling related genes in nascent and fetal HSPCs based on RNA-seq. Differentially expressed genes detected by DESeq2 were labeled. (B) SOM analysis for RNA-seq data in nascent and fetal HSPCs. (C) Expression of chd2, chd4b, smarca2, smarca4a, bptf, smarca5, and ino80 in nascent and fetal HSPCs by quantitative polymerase chain reaction (qPCR) analysis. The expression level of chd2 in nascent HSPCs was set at 1. (D) Expression of cmyb in the CHT region in control embryos and chd2, chd4b, smarca2, smarca4a, bptf, smarca5, and ino80 morphants at 3 dpf and 4 dpf by WISH. The quantification of WISH results is shown (right). Scale bars, 100 μm. Data are means ± SD (C, D). Asterisk presents statistical significance (*P < .05, **P < .01, ***P < .001). P values were calculated by a 2-tailed, unpaired Student t test. FPKM, fragments per kilobase million; mRNA, messenger RNA.
Systematical screening of chromatin remodelers controlling fetal HSPC development. (A) Scatter plots showing the expression level of chromatin remodeling related genes in nascent and fetal HSPCs based on RNA-seq. Differentially expressed genes detected by DESeq2 were labeled. (B) SOM analysis for RNA-seq data in nascent and fetal HSPCs. (C) Expression of chd2, chd4b, smarca2, smarca4a, bptf, smarca5, and ino80 in nascent and fetal HSPCs by quantitative polymerase chain reaction (qPCR) analysis. The expression level of chd2 in nascent HSPCs was set at 1. (D) Expression of cmyb in the CHT region in control embryos and chd2, chd4b, smarca2, smarca4a, bptf, smarca5, and ino80 morphants at 3 dpf and 4 dpf by WISH. The quantification of WISH results is shown (right). Scale bars, 100 μm. Data are means ± SD (C, D). Asterisk presents statistical significance (*P < .05, **P < .01, ***P < .001). P values were calculated by a 2-tailed, unpaired Student t test. FPKM, fragments per kilobase million; mRNA, messenger RNA.
To determine whether these chromatin remodelers are functionally involved in fetal HSPC development, we applied the Morpholinos knockdown strategy. WISH data showed that nascent HSPC development was not affected; however, the expression of the HSPC marker cmyb in CHT decreased after 3 dpf, indicating defective fetal HSPC development in these morphants (Figure 2D; supplemental Figure 2A). By systematically analyzing the expression pattern in ZFIN and our WISH data, we found that smarca5 was specifically expressed in CHT from 3 dpf and enriched in HSPCs (supplemental Figure 2B-C). Taken together, the spatiotemporal expression pattern of smarca5 and the hematopoietic defects in smarca5 morphants indicate that smarca5 is an essential regulator for fetal HSPC development.
smarca5 is required for fetal HSPC development
To further study the function of smarca5 in fetal HSPC development, a smarca5 mutant with a 13-bp deletion in exon 2, smarca5zko1049a, was generated using CRISPR/Cas9. The expression of smarca5 was nearly absent in smarca5zko1049a, compared with their siblings (supplemental Figure 3A-C). Morphological and gene expression experiments showed that the overall growth of smarca5 mutants is comparable with their siblings, and no developmental delay was observed (supplemental Figure 3D-E). WISH data showed that the development of nascent HSPCs was not affected, whereas fetal HSPC development was impaired, manifested as the decreased expression of cmyb in CHT (Figure 3A-B). Furthermore, the expression of the respective erythroid, myeloid, and lymphoid markers, gata1, pu1, and rag1, was all decreased in CHT or thymus at 4 dpf in smarca5zko1049a (Figure 3C). Flow cytometry and confocal imaging analysis showed that the numbers of HSPCs, thrombocytes, myeloid cells, and neutrophil cells were significantly reduced after 3 dpf in CHT in smarca5zko1049a (supplemental Figure 3F-H). Since there is no specific marker to distinguish HSCs from HSPCs in zebrafish, whether the decrease in HSPCs in smarca5zko1049a CHT occurs in HSCs or in progenitors awaits further study. In addition, it was found that Smarca5 overexpression after heat shock treatment in Tg (hsp70:smarca5-EGFP) background can efficiently rescue the defects in fetal HSPC development in smarca5zko1049a (Figure 3D-F), suggesting that smarca5 loss is responsible for fetal HSPC defects in these mutants.
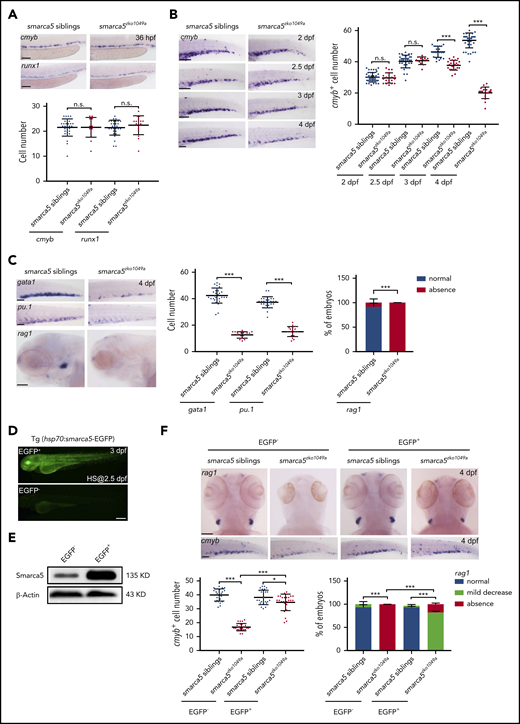
Fetal HSPC development is impaired in smarca5 mutants. (A) Expression of cmyb and runx1 in the AGM region in smarca5zko1049a and their siblings at 36 hpf by WISH. The quantification of WISH results is shown (bottom). Scale bars, 100 μm. (B) Expression of cmyb in the CHT region in smarca5zko1049a and their siblings from 2 dpf to 4 dpf by WISH. The quantification of WISH results is shown (right). Scale bars, 100 μm. (C) Expression of erythroid marker gata1, myeloid marker pu.1, and lymphoid marker rag1 in the CHT or thymus region in smarca5zko1049a and their siblings at 4 dpf by WISH. The quantification of WISH results is shown (right). Scale bars, 100 μm. (D) The imaging of enhanced green fluorescent protein (EGFP) fluorescence in Tg (hsp70:smarca5-EGFP) embryos at 3 dpf. Heat shock was performed at 2.5 dpf. Scale bars, 200 μm. (E) Protein level of Smarca5 in EGFP− and EGFP+ embryos in (D) at 3 dpf by western blot. Heat shock was performed at 2.5 dpf. (F) Expression of cmyb in the CHT region and rag1 in the thymus region in smarca5zko1049a and their siblings, with or without Smarca5 overexpression from 2 dpf to 3 dpf by WISH. The quantification of WISH results is shown (bottom). Scale bars, 100 μm. Data are means ± SD (A-C, F). Asterisk presents statistical significance (*P < .05, ***P < .001; n.s., not significant). P values were calculated by a 2-tailed, unpaired Student t test. HS, heat shock.
Fetal HSPC development is impaired in smarca5 mutants. (A) Expression of cmyb and runx1 in the AGM region in smarca5zko1049a and their siblings at 36 hpf by WISH. The quantification of WISH results is shown (bottom). Scale bars, 100 μm. (B) Expression of cmyb in the CHT region in smarca5zko1049a and their siblings from 2 dpf to 4 dpf by WISH. The quantification of WISH results is shown (right). Scale bars, 100 μm. (C) Expression of erythroid marker gata1, myeloid marker pu.1, and lymphoid marker rag1 in the CHT or thymus region in smarca5zko1049a and their siblings at 4 dpf by WISH. The quantification of WISH results is shown (right). Scale bars, 100 μm. (D) The imaging of enhanced green fluorescent protein (EGFP) fluorescence in Tg (hsp70:smarca5-EGFP) embryos at 3 dpf. Heat shock was performed at 2.5 dpf. Scale bars, 200 μm. (E) Protein level of Smarca5 in EGFP− and EGFP+ embryos in (D) at 3 dpf by western blot. Heat shock was performed at 2.5 dpf. (F) Expression of cmyb in the CHT region and rag1 in the thymus region in smarca5zko1049a and their siblings, with or without Smarca5 overexpression from 2 dpf to 3 dpf by WISH. The quantification of WISH results is shown (bottom). Scale bars, 100 μm. Data are means ± SD (A-C, F). Asterisk presents statistical significance (*P < .05, ***P < .001; n.s., not significant). P values were calculated by a 2-tailed, unpaired Student t test. HS, heat shock.
To determine whether smarca5 loss would lead to any endothelial defects in CHT, we first detected the morphological feature of CHT vasculature in smarca5 mutants and their siblings. Confocal imaging by using kdrl:mCherry showed that there is no discernible difference in the CHT vasculature between smarca5 mutants and their siblings at 3 dpf and 4 dpf (supplemental Figure 3I). To explore whether the CHT niche in smarca5 mutants has any influence on HSPC development, we generated parabiotic embryo pairs31,32 with smarca5 mutants and siblings in Tg (CD41:GFP) or Tg (gata1:dsRed) background (supplemental Figure 3J). A normal number of CD41:GFP+ cells from smarca5 siblings was observed in the smarca5 mutant CHT region. However, far fewer CD41:GFP+ cells from smarca5 mutants were observed in the CHT of smarca5 mutants and siblings (supplemental Figure 3K). Together, these results demonstrated that the HSPC defects in smarca5 mutants is mainly attributed to HSPC intrinsic defects, but not influenced by the CHT niche.
We further evaluated the cell proliferation, cell-cycle progression, and cell death status in smarca5zko1049a and their siblings. The percentage of 5-bromodeoxyuridine+cmyb+ cells in cmyb+ cells was decreased significantly (P < .001) in smarca5zko1049a, and the smarca5 mutation induced a mild delay in cell-cycle progression in S phase in HSPCs compared with their sibling HSPCs (supplemental Figure 4A-B). The number of apoptotic cells was increased significantly (P < .001) in CHT (supplemental Figure 4C), after 3 dpf in smarca5zko1049a, compared with their siblings. A previous study found that the p53 signaling was upregulated in smarca5-deficient HSPCs.33 In smarca5zko1049a, we also observed the increased expression of p53 and p21 (supplemental Figure 4D). However, knockdown or knockout p53 cannot rescue the hematopoietic defects in smarca5zko1049a, suggesting that the hematopoietic phenotype in smarca5zko1049a is largely p53 independent (supplemental Figure 4E). However, treatment of embryos with pan-caspase inhibitor (Z-VAD-FMK) resulted in the decrease of TUNEL signals and an efficient rescue of cmyb expression in CHT in smarca5zko1049a (supplemental Figure 4F-G), indicating that smarca5 deletion leads to a caspase-dependent apoptosis in HSPCs. Altogether, our results show that loss of smarca5 impairs fetal HSPC development in CHT after 3 dpf.
Loss of Smarca5 disrupts chromatin accessibility
To determine whether Smarca5 can modulate site-specific chromatin accessibility, we performed ATAC-seq with fetal HSPCs in smarca5zko1049a and their siblings (supplemental Figure 5A). Principal component analysis showed that the smarca5 sibling HSPCs tend to group closely with HSPCs in wild-type, but not with smarca5-deficient HSPCs (supplemental Figure 5B). By calculating the number of ATAC-seq peaks, we observed the changes in chromatin accessibility across the genome upon smarca5 deletion, with the number of peaks changed from 27 438 to 13 375. In brief, 12 201 peaks remained unaltered between HSPCs in smarca5zko1049a and their siblings; 218 peaks were specifically accessible in mutants, whereas 4150 peaks were specifically accessible in siblings (Figure 4A). GO analysis showed that in smarca5-deficient HSPCs, the enrichment of hematopoiesis-related genes with accessible chromatin was decreased (supplemental Figure 5C). The genes with decreased chromatin accessibility at promoters after smarca5 deletion were enriched for the terms of “myeloid cell homeostasis,” “myeloid cell differentiation,” “homeostasis of number of cells,” “erythrocyte homeostasis,” and “erythrocyte differentiation” (Figure 4B), indicating that smarca5 deletion alters the chromatin accessibility of genes related to the maintenance of HSPC expansion and differentiation.
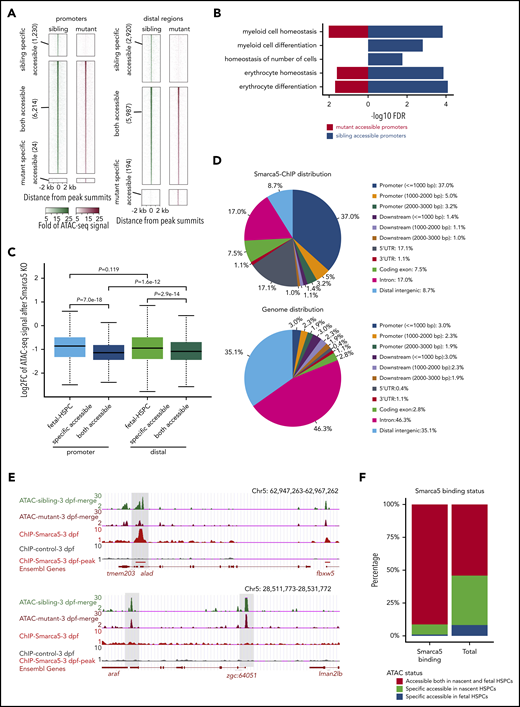
Deletion of smarca5 leads to the decrease of chromatin accessibility. (A) Heat maps showing ATAC-seq signals around ATAC-seq peaks in sibling and mutant at promoters and distal regulatory regions. Peaks were classified into sibling-specific accessible, both accessible, and mutant-specific accessible. In each group, peaks are ranked according to ATAC-seq signals. (B) GO analysis of genes with accessible promoter regions in HSPCs in smarca5zko1049a and their siblings. (C) Box plot showing the log2-transformed fold change of ATAC-seq signals at fetal HSPC-specific and both accessible regions at promoters and distal regulatory regions after smarca5 knockout (KO). smarca5 KO leads to significant decrease of ATAC-seq signals especially at both accessible promoter regions. P values were calculated by a 2-tailed Wilcoxon test. (D) Pie charts drew by CEAS56 showing the distribution of Smarca5-ChIP-seq peaks (top) and genome distribution background (bottom) across the genome. (E) The genome browser views showing ATAC-seq signals of genes with (top) or without (bottom) Smarca5 binding. (F) Bar plot showing the proportion of Smarca5 binding sites distributed on both accessible chromatin regions in nascent and fetal HSPCs, specific accessible regions in nascent or fetal HSPCs compared with background (total). UTR, untranslated region.
Deletion of smarca5 leads to the decrease of chromatin accessibility. (A) Heat maps showing ATAC-seq signals around ATAC-seq peaks in sibling and mutant at promoters and distal regulatory regions. Peaks were classified into sibling-specific accessible, both accessible, and mutant-specific accessible. In each group, peaks are ranked according to ATAC-seq signals. (B) GO analysis of genes with accessible promoter regions in HSPCs in smarca5zko1049a and their siblings. (C) Box plot showing the log2-transformed fold change of ATAC-seq signals at fetal HSPC-specific and both accessible regions at promoters and distal regulatory regions after smarca5 knockout (KO). smarca5 KO leads to significant decrease of ATAC-seq signals especially at both accessible promoter regions. P values were calculated by a 2-tailed Wilcoxon test. (D) Pie charts drew by CEAS56 showing the distribution of Smarca5-ChIP-seq peaks (top) and genome distribution background (bottom) across the genome. (E) The genome browser views showing ATAC-seq signals of genes with (top) or without (bottom) Smarca5 binding. (F) Bar plot showing the proportion of Smarca5 binding sites distributed on both accessible chromatin regions in nascent and fetal HSPCs, specific accessible regions in nascent or fetal HSPCs compared with background (total). UTR, untranslated region.
In addition, we found a significant decrease in chromatin accessibility at conserved accessible regions compared with fetal HSPC-specific accessible regions upon smarca5 deletion. Of note, there was a greater decrease in the level of chromatin accessibility in promoters than in distal regulatory regions (Figure 4C). To further investigate the downstream targets of Smarca5, we performed Smarca5 ChIP-seq (supplemental Figure 5D-E). As shown by the pie charts (Figure 4D), Smarca5 tends to bind to the promoters. Representative examples showed that upon smarca5 deletion, a decrease in chromatin accessibility was observed in Smarca5 binding regions but not at the non–Smarca5-binding sites (Figure 4E), indicating the specificity of Smarca5 in regulating chromatin accessibility. Moreover, we observed that Smarca5-binding sites were mainly distributed in the conserved accessible regions between nascent and fetal HSPCs (Figure 4F), and smarca5 deletion led to a significant decrease of chromatin accessibility in these regions (supplemental Figure 5F), indicating the functional role of Smarca5 in regulating chromatin accessibility at promoters of hematopoiesis-related genes during HSPC development.
To further study the change of transcriptome in HSPCs in smarca5zko1049a, RNA-seq was performed with HSPCs in CHT at 3 dpf in smarca5zko1049a and their siblings (supplemental Figure 5A). Using DESeq2,34 we identified 308 and 705 genes with higher expression in either sibling or mutant HSPCs, respectively. Among these genes, many hematopoiesis-related genes, such as gata2a, alas2, runx3, and irf4b, displayed decreased expression in smarca5zko1049a (supplemental Figure 5G). Moreover, expression of genes related to cell cycle and cell survival, including p53, cdkn1a, mcm3, baxa, gadd45aa, and caspb, was altered in smarca5zko1049a (supplemental Figure 5G), which is consistent with the observed defects in cell proliferation and cell survival of HSPCs in smarca5zko1049a (supplemental Figure 4A-C).
Taken together, our results suggest that during fetal HSPC development, Smarca5 is required for the maintenance of chromatin accessibility at promoters of hematopoiesis-related genes, which are responsible for the maintenance of HSPC expansion and differentiation.
Smarca5 interacts with Ncl to facilitate chromatin remodeling
To identify the potential interacting proteins of Smarca5 in fetal HSPC development, we performed mass spectrometry (MS) analysis of Flag-Smarca5 pull-down proteins (supplemental Figure 6A-B). We found that Ybx1 and Ncl were on the top list, and the corresponding encoding genes ybx1 and ncl are coexpressed with smarca5 in CHT (Figure 5A-B). In addition, by analyzing our previous bulk RNA-seq data of HSPCs,27 we found that ncl is highly expressed in HSPCs at 3 dpf (supplemental Figure 6C). Moreover, CoIP analysis showed that the interaction between Smarca5 and Ncl is stronger than that between Smarca5 and Ybx1 (Figure 5C). Together, these results indicate that Smarca5 and Ncl may work together to facilitate the development of fetal HSPCs.
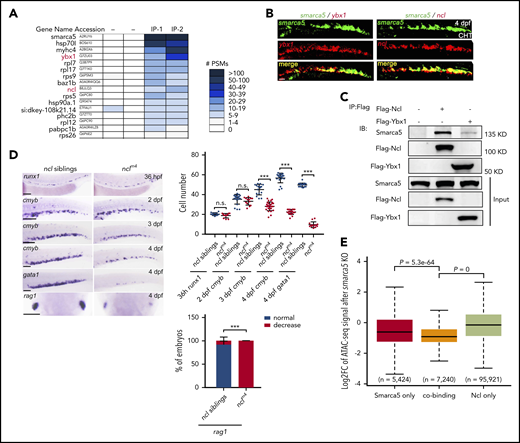
Ncl collaborates with Smarca5 to facilitate chromatin remodeling. (A) Analysis of the interaction proteins with Smarca5 using MS. Heat maps showing the number of peptide spectrum match (PSM) identified by MS for proteins after immunoprecipitation of Flag-Smarca5 from the CHT region of zebrafish embryos at 3 dpf. (B) Double-fluorescence in situ hybridization analysis showing the coexpression between ybx1, ncl with smarca5 in CHT. Scale bars, 50 μm. (C) Zebrafish embryos were injected with the indicated plasmids at 1-cell stage and CoIP was performed at 3 dpf, immunoblot analysis with the indicated antibodies showing the interaction between Ybx1, Ncl, and Smarca5. (D) Expression of runx1, cmyb, gata1, and rag1 in the AGM, CHT, or thymus regions in nclm4 and their siblings from 36 hpf to 4 dpf by WISH. The quantification of WISH results is shown (right). Scale bars, 100 μm. (E) Box plot showing the log2-transformed fold change of ATAC-seq signals at after smarca5 knockout at Smarca5 only, cobinding, and Ncl only regions. P values were calculated by a 2-tailed Wilcoxon test. Data are mean ± SD. (D) Asterisk presents statistical significance (***P < .001; n.s., not significant). P values were calculated by a 2-tailed, unpaired Student t test.
Ncl collaborates with Smarca5 to facilitate chromatin remodeling. (A) Analysis of the interaction proteins with Smarca5 using MS. Heat maps showing the number of peptide spectrum match (PSM) identified by MS for proteins after immunoprecipitation of Flag-Smarca5 from the CHT region of zebrafish embryos at 3 dpf. (B) Double-fluorescence in situ hybridization analysis showing the coexpression between ybx1, ncl with smarca5 in CHT. Scale bars, 50 μm. (C) Zebrafish embryos were injected with the indicated plasmids at 1-cell stage and CoIP was performed at 3 dpf, immunoblot analysis with the indicated antibodies showing the interaction between Ybx1, Ncl, and Smarca5. (D) Expression of runx1, cmyb, gata1, and rag1 in the AGM, CHT, or thymus regions in nclm4 and their siblings from 36 hpf to 4 dpf by WISH. The quantification of WISH results is shown (right). Scale bars, 100 μm. (E) Box plot showing the log2-transformed fold change of ATAC-seq signals at after smarca5 knockout at Smarca5 only, cobinding, and Ncl only regions. P values were calculated by a 2-tailed Wilcoxon test. Data are mean ± SD. (D) Asterisk presents statistical significance (***P < .001; n.s., not significant). P values were calculated by a 2-tailed, unpaired Student t test.
To further investigate the function of ncl in fetal HSPC development, a ncl mutant line with a 4-bp deletion in exon 2, nclm4, was generated using CRISPR/Cas9 (supplemental Figure 6D). The expression level of ncl was nearly absent in nclm4 compared with their siblings (supplemental Figure 6E-F). In nclm4, runx1 expression in 36 hpf AGM was normal, indicating that the development of nascent HSPCs was not affected. However, the development of fetal HSPCs was impaired, with decreased expression of cmyb in CHT after 3 dpf (Figure 5D). Moreover, the number of HSPCs in CHT was decreased significantly (P < .001) in nclm4 compared with their siblings (supplemental Figure 6G-H), indicating the essential role of ncl in fetal HSPC development.
To examine the impact of Ncl on Smarca5-mediated chromatin remodeling, we performed Ncl ChIP-seq. There was a significant overlap (P = 0) between the genes bound by Smarca5 (6077) and Ncl (13 757) in their promoters (supplemental Figure 6I). Moreover, the accessibility of 2 factors cobinding regions showed a significantly decrease in smarca5zko1049a, compared with those bound by Smarca5 or Ncl alone (Figure 5E). Those results indicate that Ncl plays a role in facilitating Smarca5-mediated chromatin remodeling.
Genomic binding of hematopoietic TFs depends on Smarca5-mediated chromatin remodeling
A recent study has demonstrated the critical role of SMARCA5 in mediating the correct genomic binding of TFs in mouse embryonic stem cells.35 However, it is largely unknown whether Smarca5 can affect the binding of hematopoietic TFs during fetal HSPC development. We thus screened the motifs enriched in sibling HSPC-specific accessible chromatin regions. The hematopoietic TF DNA-binding motifs, such as Klf1 and Spi1, were included on the top list, indicating that Smarca5 could facilitate genomic binding of TFs that are associated with the maintenance and differentiation programs of HSPCs (Figure 6A). Moreover, the comparable expression level of klf1, spi1b, yy1a, yy1b, and spic between smarca5zko1049a and their siblings excluded the secondary effects from decreased expression level of these factors (supplemental Figure 7). Next, we selected target genes of these TFs with a decrease in chromatin accessibility in smarca5zko1049a for further expression analysis. qPCR results validated the significantly decreased expression of bcl11ab, crtc3, chaf1b, and nfe2l1b in smarca5zko1049a (Figure 6B).
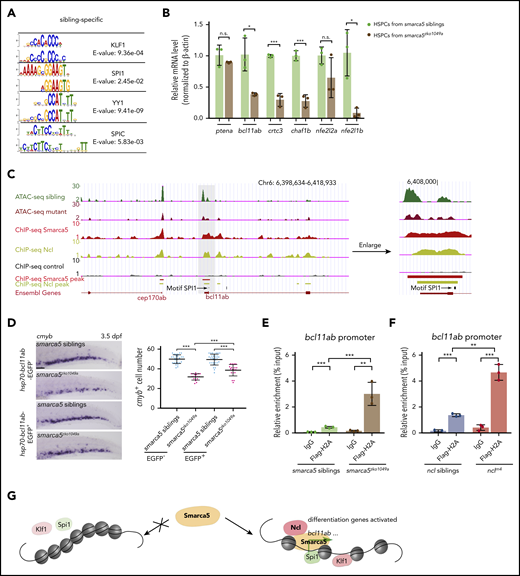
Smarca5 deletion impairs the genomic binding of hematopoietic TFs. (A) Motif scan results at sites with lost accessibility after smarca5 deletion. For each presented pair, the top one is the motif from JASPAR database; the bottom one is the scanned motif. (B) qPCR analysis showing the expression of ptena, bcl11ab, crtc3, chaf1b, nfe2l2a, and nfe2l1b in HSPCs from smarca5zko1049a and their siblings. (C) The genome browser views showing the track of Smarca5 and Ncl occupancy in bcl11ab promoter with ATAC-seq peaks. The gray box indicates the change of ATAC-seq peaks after smarca5 deletion. The location of a predicted Spi1 binding site is marked by a black line. (D) Expression of cmyb in the CHT region in smarca5zko1049a and their siblings at 3.5 dpf, with or without Bcl11ab overexpression from 2 dpf to 3 dpf by WISH. The quantification of WISH results is shown (right). Scale bars, 100 μm. (E) ChIP-qPCR analysis of H2A occupancy compared with immunoglobulin G (IgG) in the promoter of bcl11ab in smarca5zko1049a and their siblings. (F) ChIP-qPCR analysis of H2A occupancy compared with IgG in the promoter of bcl11ab in nclm4 and their siblings. (G) A model shows that Smarca5-mediated chromatin accessibility programming is responsible for fetal HSPC development. The chromatin remodeler smarca5 plays an important role in regulating chromatin accessibility to facilitate the binding of hematopoietic TFs, such as Klf1 and Spi1. Moreover, Ncl is identified to collaborate with Smarca5 to facilitate efficient chromatin remodeling. Data are means ± SD (B,D-F). Asterisk presents statistical significance (*P < .05, **P < .01, ***P < .001; n.s., not significant). P values were calculated by a 2-tailed, unpaired Student t test.
Smarca5 deletion impairs the genomic binding of hematopoietic TFs. (A) Motif scan results at sites with lost accessibility after smarca5 deletion. For each presented pair, the top one is the motif from JASPAR database; the bottom one is the scanned motif. (B) qPCR analysis showing the expression of ptena, bcl11ab, crtc3, chaf1b, nfe2l2a, and nfe2l1b in HSPCs from smarca5zko1049a and their siblings. (C) The genome browser views showing the track of Smarca5 and Ncl occupancy in bcl11ab promoter with ATAC-seq peaks. The gray box indicates the change of ATAC-seq peaks after smarca5 deletion. The location of a predicted Spi1 binding site is marked by a black line. (D) Expression of cmyb in the CHT region in smarca5zko1049a and their siblings at 3.5 dpf, with or without Bcl11ab overexpression from 2 dpf to 3 dpf by WISH. The quantification of WISH results is shown (right). Scale bars, 100 μm. (E) ChIP-qPCR analysis of H2A occupancy compared with immunoglobulin G (IgG) in the promoter of bcl11ab in smarca5zko1049a and their siblings. (F) ChIP-qPCR analysis of H2A occupancy compared with IgG in the promoter of bcl11ab in nclm4 and their siblings. (G) A model shows that Smarca5-mediated chromatin accessibility programming is responsible for fetal HSPC development. The chromatin remodeler smarca5 plays an important role in regulating chromatin accessibility to facilitate the binding of hematopoietic TFs, such as Klf1 and Spi1. Moreover, Ncl is identified to collaborate with Smarca5 to facilitate efficient chromatin remodeling. Data are means ± SD (B,D-F). Asterisk presents statistical significance (*P < .05, **P < .01, ***P < .001; n.s., not significant). P values were calculated by a 2-tailed, unpaired Student t test.
Based on the screening results, the chromatin accessibility at bcl11ab promoters, which contains sites also occupied by Ncl, was decreased in smarca5zko1049a (Figure 6C). Given that Bcl11ab is required for human B lymphopoiesis and hemoglobin switching36-38 and mouse FL HSPC development,39 we thus generated an hsp70:bcl11ab-EGFP transgenic line in smarca5zko1049a background. We found that overexpression of bcl11ab after 2 dpf can efficiently rescue the defects of fetal HSPC development in smarca5zko1049a (Figure 6D), suggesting that bcl11ab acts as a downstream target of Smarca5.
Previous studies suggested that Ncl influenced the nucleosome sliding by promoting H2A/H2B decomposition in vitro,40 and we questioned whether Ncl also participates in H2A/H2B decomposition in fetal HSPC development. Flag-H2A ChIP-qPCR analyses showed that H2A signals were more enriched at the promoter of bcl11ab in smarca5zko1049a or nclm4 compared with their siblings (Figure 6E-F), indicating that the abnormal decomposition of H2A/H2B possibly leads to the decreased expression of bcl11ab. These results show that the co-remodeler Ncl collaborates with Smarca5 to promote chromatin remodeling, possibly through regulating H2A/H2B deposition. Overall, Smarca5, in collaboration with a co-remodeler Ncl, can mediate chromatin accessibility to promote genomic binding of hematopoietic TFs, which further activate the expression of target genes during fetal HSPC development.
A conserved role of chromatin remodelers in mammalian fetal HSPCs
In mice, nascent HSCs acquire definitive HSC characteristics only when they colonize the FL.9,10 To investigate whether the regulatory mechanism of chromatin remodelers in fetal HSPCs identified above is also conserved in mammals, we first examined the expression pattern of chromatin remodelers in primary human FL cells by analyzing the single cell RNA-seq data in human FL.41 Uniform manifold approximation and projection assay showed that the expression of many chromatin remodelers, including Smarca5, was highly enriched in human FL hematopoietic cells, especially in HSC and hematopoietic progenitor clusters (supplemental Figure 8A). These results suggest the conserved expression pattern of chromatin remodelers in vertebrate fetal hematopoiesis, from fish to humans.
To further identify the conserved function of Smarca5 in mammalian fetal HSPC development, we knocked down endogenous Smarca5 expression in mouse E14.5 FL lineage-sca1+c-Kit+ cells using Smarca5 short interfering RNA (supplemental Figure 8B-C) and performed the colony-forming unit cell assay. The results showed that the formation ability of granulocyte, erythrocyte, macrophage, and megakaryocyte and granulocyte-macrophage (GM) colonies of FL LSK cells was significantly impaired (P < .05) upon Smarca5 knockdown (supplemental Figure 8D). These data indicate the specific functional role of Smarca5 in fetal HSPC development, which is similar to the observed phenotype of Smarca5 mutants in mice.33 Collectively, our findings support a well-conserved role of chromatin remodelers during fetal HSPC development in mammals.
Discussion
Our ATAC-seq and RNA-seq analyses show that the development of HSPCs from nascent to fetal states undergoes dynamic changes in chromatin accessibility and transcriptome features. We then screen and demonstrate that smarca5 is essential for fetal HSPC development by maintaining the accessible chromatin regions to facilitate the genomic binding of hematopoietic TFs. Moreover, we identify a co-remodeler, Ncl, which collaborates with Smarca5 to ensure efficient chromatin remodeling in vivo. A model is thus proposed to indicate the dynamics of chromatin accessibility and transcriptome features between nascent and fetal HSPCs and that Smarca5-mediated epigenetic programming is critical for fetal HSPC development (Figure 6G). However, smarca5 overexpression is not sufficient to drive transcriptome transition from nascent to fetal HSPC state (data not shown), indicating the requirement but not sufficient role of smarca5 in fetal HSPC development.
Chromatin accessibility has been extensively studied in pluripotent stem cell reprogramming in vitro.42-44 These studies show that stem cells, including human embryonic stem cells, possess more open chromatin state compared with the differentiated cells and that cell fate specification is accompanied by a transition of chromatin organization from an accessible state to a restricted state. Chromatin state transition is also critical for the maternal to zygotic transition in early embryo development. In this process, maternal chromatin remodeler smarca2 plays an important role in inhibiting chromatin condensation before maternal to zygotic transition, while this regulation by smarca2 is gradually erased due to the degradation of smarca2 mediated by microRNA miR-430 after the start of zygotic transcription.45 These studies demonstrate the critical role of chromatin accessibility dynamics during different biological processes. In zebrafish, a previous study identified multiple chromatin remodelers, which are functionally required in primitive and definitive hematopoiesis, such as smarca1, chrac1, and chd7 in primitive erythroid progenitors and smarcc1a and chd7 in HSPC production in AGM.46 However, the function of these factors at later hematopoietic developmental stages (ie, fetal HSPCs) is not examined. Here, we focus on the dynamic chromatin accessibility in HSPC transition from nascent to fetal state and find a selected group of chromatin remodelers are dynamically expressed and functionally required during this process. We reason that these chromatin remodelers fine tune the accessible chromatin regions and facilitate nascent HSPCs to acquire robust definitive characteristics upon transitioning into fetal HSPCs. Notably, aberrant expression of chromatin remodelers was observed in acute myeloid leukemia,47 suggesting the functional role of chromatin remodelers in regulating physiology-to-pathology transition. Elucidation of the molecular mechanism of chromatin remodelers would help understand the contribution of aberrant expression of chromatin remodelers to the progression of hematological diseases and devise personalized therapeutic strategy for effective treatment.
As the major nucleolar protein, Ncl involvement in ribosome RNA biogenesis has been well established.48,49 Moreover, Ncl is capable of decondensing chromatin for transcription.50 Previous studies have demonstrated that Ncl could regulate gene transcription in hematopoietic and cardiac systems.51-54 Furthermore, Ncl can act as a co-remodeler of chromatin remodeling, by collaborating with the SWI/SNF complex and the ATP-utilizing chromatin assembly and remodeling factor complex, and Ncl could obviously enhance the nucleosome sliding activity of SWI/SNF in vitro.40,55 In our study, we find that Ncl functions as a co-remodeler of Smarca5 in hematopoietic system in vivo, reflecting the multifunctional role of Ncl in chromatin remodeling. In addition, although the hematopoietic phenotype in nclm4 mimics that in smarca5zko1049a, the severity of hematopoietic defects in the former is stronger than that in the latter, reflected by the decreased expression of cmyb in CHT early at 2 dpf, which is not observed in smarca5zko1049a at the same time point. These data suggest that additional mechanism mediated by ncl may exist in hematopoietic system, possibly by its roles in regulating gene expression or ribosome biogenesis.
In summary, we have demonstrated epigenetic programming is critical for fetal HSPC development in vertebrates. These findings raise the possibility that the modulation of chromatin remodelers, TFs and microenvironmental signals,27 can direct cell-state transition and thus provides implications for the improvement of protocols to generate functional HSPCs in vitro.
The raw ATAC-seqA-seq and ChIP-seq data reported in this paper have been deposited in the Genome Sequence Archive in the National Genomics Data Center, Beijing Institute of Genomics (Chinese National Center for Bioinformation), Chinese Academy of Sciences (http://bigd.big.ac.cn/gsa), under accession number CRA001858 and are publicly accessible at https://bigd.big.ac.cn/gsa. The shared URL for reviewers is http://bigd.big.ac.cn/gsa/s/QRPx1S00.
The online version of this article contains a data supplement.
The publication costs of this article were defrayed in part by page charge payment. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734.
Acknowledgments
The authors thank Bertie Gottgens for critical reading of the manuscript.
This work was supported by grants from the National Key Research and Development Program of China (2018YFA0800200, 2018YFA0801000, 2016YFA0100500, 2017YFA0102600, 2016YFA0100400), the Strategic Priority Research Program of the Chinese Academy of Sciences, (XDA16010207), and the National Natural Science Foundation of China (31830061, 31425016, and 81530004). This study was supported by the State Key Laboratory of Membrane Biology, China and Translational Medical Center for Stem Cell Therapy and Institute for Regenerative Medicine, China.
Authorship
Contribution: Y.D. performed most of the experiments with help from D.M., G.L., Y.X., Yifan Zhang, L.W., and J.H.; W.W. performed the bioinformatics analysis under the guidance of Yong Zhang; Z.K. performed the SOM analysis; Y.D., W.W., Yong Zhang, and F.L. conceived the project, analyzed the data, and wrote the paper; and all authors read and approved the final manuscript.
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Correspondence: Feng Liu, State Key Laboratory of Membrane Biology, Institute of Zoology, Chinese Academy of Sciences, Beijing, 100101, China; e-mail: liuf@ioz.ac.cn; and Yong Zhang, School of Life Science and Technology, Tongji University, Shanghai 200092, China; e-mail: yzhang@tongji.edu.cn.

This feature is available to Subscribers Only
Sign In or Create an Account Close Modal